11.1 The Cell Membrane
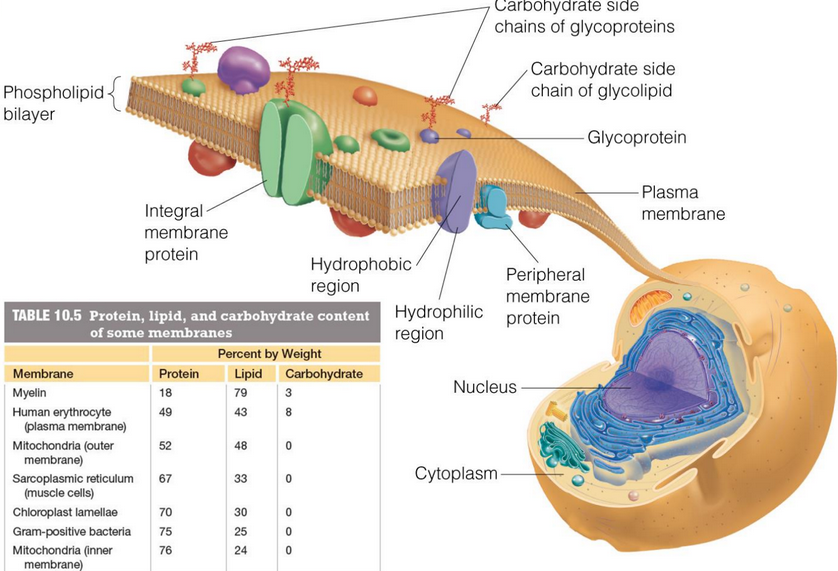
Figure 11.2: Features of the Cell Membrane
Some features of the cell membrane are shown in figure 11.2.
11.1.1 History of the cell membrane
In 1935, scientists Hugh Davson and James Danielli portrayed the cell membrane as a “sandwich”:
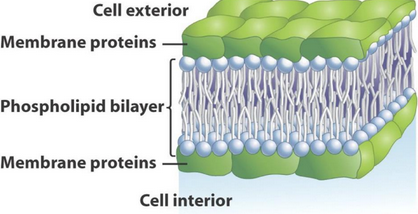
Figure 11.3: Sandwich Model of the Cell Membrane
In this model (see figure 11.3), the phospholipid bilayer of the membrane lies between two layers of globular proteins. However, the placement of these globular proteins have hydrophilic and hydrophobic regions!
In 1972, scientists J. Singer and G. Nicholson revised the model in figure 11.3 in a simple, yet profound way:
They proposed that the globular proteins in figure 11.3 were instead inserted into the lipid bilayer; the nonpolar segments of proteins are in contact with the nonpolar regions of the interior of the bilayer and the polar portions protruding out of the cell membrane:
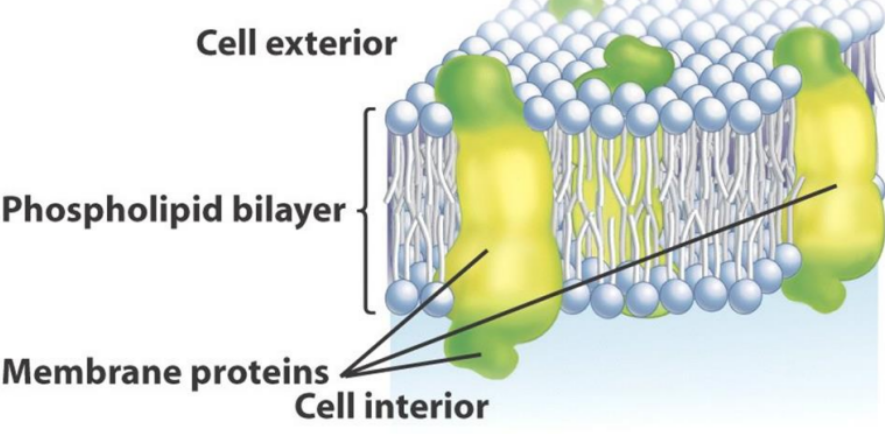
Figure 11.4: Singer and Nicholson’s Model of the Cell Membrane
The model in 11.4 is called the fluid mosaic model.
11.1.2 Lipid asymmetry
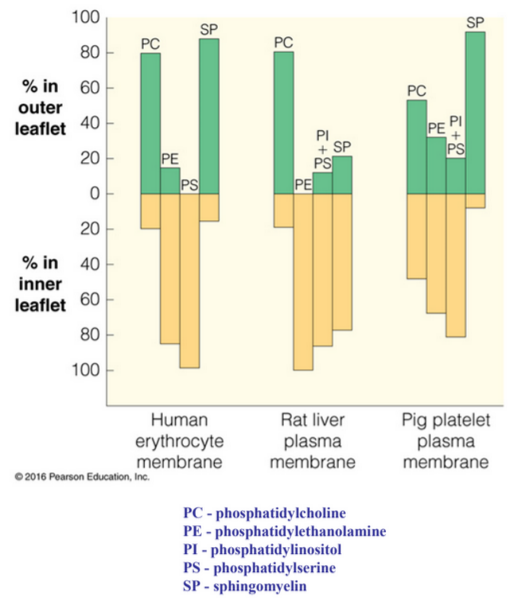
Figure 11.5: Distribution of Lipids Inside and Outside the Cell Membrane
Glycolipids on the outer portion of the cell membrane likely serve as receptors for extracellular signals.
Phosphatidylserine on the inner leaflet of the membrane has a net negative charge at physiological pH, hence making it an ideal candidate for binding to positively-charged lysine and arginine residues.
However, the appearance of phosphatidylserine on the outer membranes of aging lymphocytes marks the cells for destruction by macrophages. On the surface of platelets, phosphatidylserines cause blood coagulation.
11.1.3 Components of cell membranes
Figure 11.6: Components of the Cell Membrane
Figure 11.6 displays several components of the cell membrane.
11.1.4 Interior protein networks
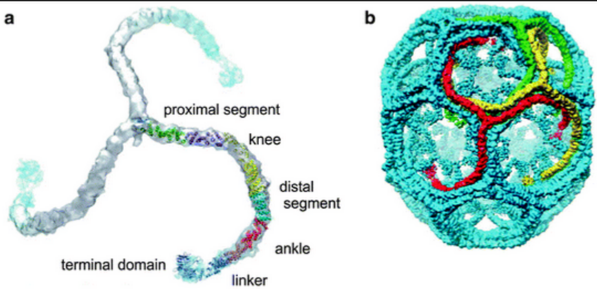
Figure 11.7: An Interior Protein Network
The interior protein network clathrin (see figure 11.7) is the main constituent of coated pits and vesicles. It is a soluble, cytosolic protein that binds lipids directly and hence, is recruited to the membrane by a specialized set of proteins called adaptors to assemble into coated pits and vesicles.
Approximately 100 clathrin triskelion units are present in an assembled coated vesicle (also see figure 11.7).