5.2 Myoglobin and Hemoglobin
Myoglobin and hemoglobin are heme-carrying proteins that are able to bind molecular oxygen and carry out functions for oxygen storage and transport respectively.
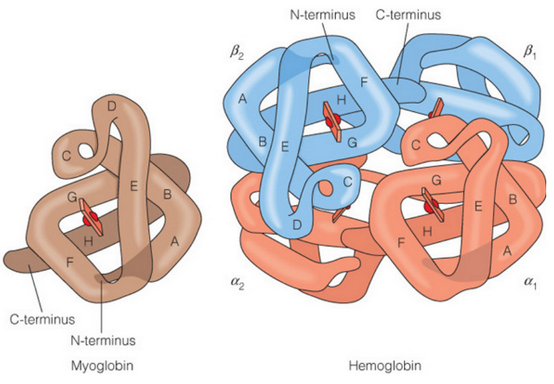
Figure 5.3: Side-By-Side Comparison of Hemoglobin and Myoglobin
Like seen in figure 5.3, myoglobin is a single subunit protein. Hemeglobin has four subunits: two \(\alpha\) subunits and two \(\beta\) subunits. Both the \(\alpha\) and the \(\beta\) subunits are similar.
Myoglobin and hemoglobin are very structurally similar - this is because they arose from a common ancestral gene (via gene duplication), only to diverge and assume specialized roles in oxygen storage and transport.
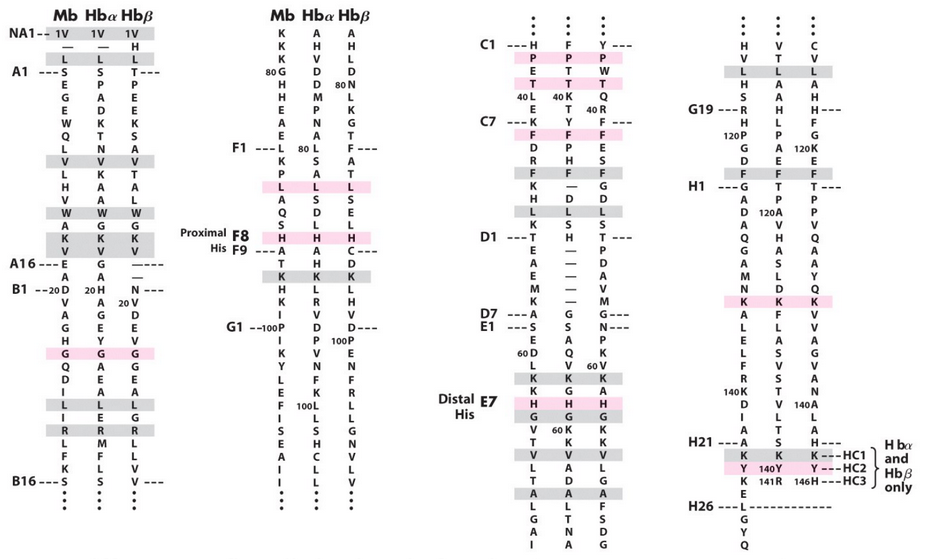
Figure 5.4: Amino Acid Sequences of Hemoglobin and Myoglobin
Figure 5.4 depicts the amino acid sequence of hemoglobin and myoglobin. The differences in amino acid sequences all reflect a distinction in function. A major part of this distinction is a tetrameric form for hemoglobin.
5.2.1 O2 binding in the globins
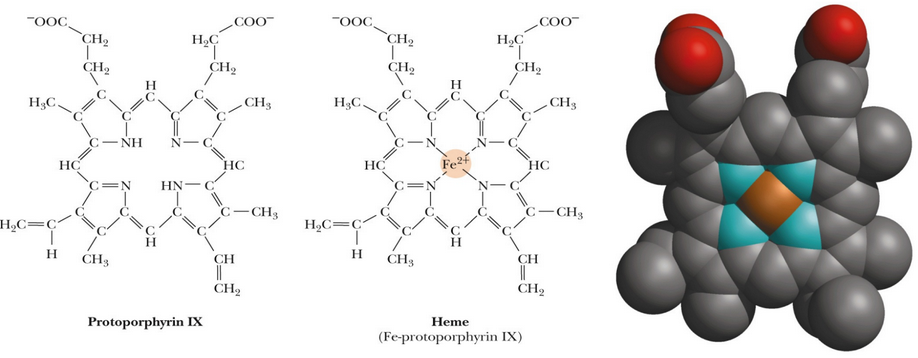
Figure 5.5: Side-by-Side Comparison of Heme and Protoporphyrin
Many proteins have evolved to use metal ions for their activity; in these cases, the special chemistry of metals is necessary for binding, catalytic, and structural functions.
In other systems (e.g., prosthetic groups), protoporphyrin IX (see figure 5.5) is necessary to modulate the properties of the bound metal. A heme-protein active site suits the special requirements associated with binding O2.
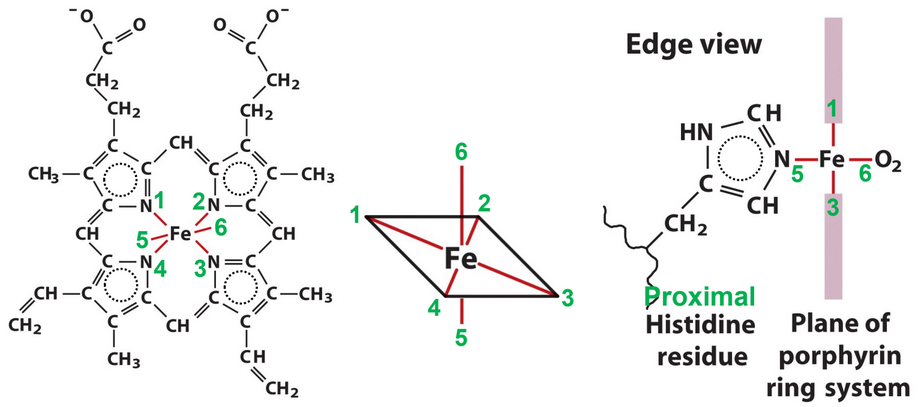
Figure 5.6: Iron in Hemoglobin
The transition metal ion Fe2+ can form covalent bonds with polarizable ligands - in particular, aromatic ring nitrogen groups like histidine and porphyrin ring systems. Fe2+ assumes an octahedral coordination geometry (see figure 5.6) with six ligands: four porphyrin N groups, a proximal His ligand, and a sixth location for binding O2.
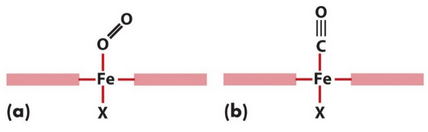
Figure 5.7: Carbon Monoxide versus Oxygen Binding
Carbon monoxide (i.e., CO) binds to heme in a straight fashion (i.e., (a) of figure 5.7) while O2 binds to heme in a bent fashion (i.e., (b) of figure 5.7).
Also note that O2 binding in hemoglobin stabilized by an additional hydrogen bond from a distal histidine.
5.2.2 O2 storage and transport problem
Myoglobin has a very high affinity for O2 at all partial pressures of O2, hence making it ideal for an O2 storage protein. However, hemoglobin is different: it binds to O2 in the lungs and releases it into body tissues.
The partial pressure of O2 in the lungs is about 100 mm Hg - hemoglobin becomes saturated here. However, the partial pressure of O2 in the capilaries is about 30 mm Hg - O2 is hence released from here.
To achieve the latter, the binding of O2 to hemoglobin is coorperative: when O2 binds to one subunit, there are structural changes that make binding to the other subunits more favorable.
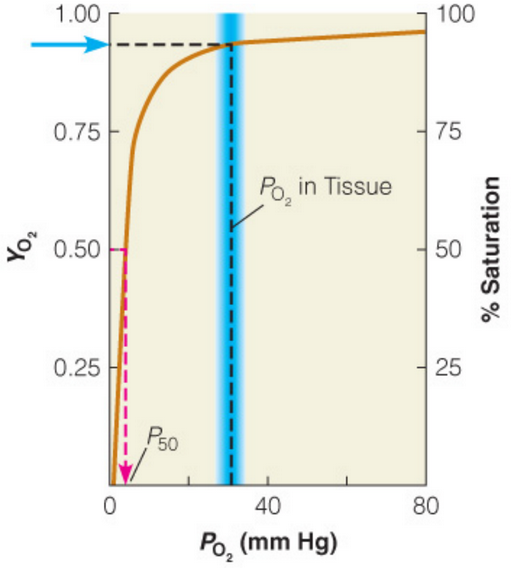
Figure 5.8: Oxygen Binding in Myoglobin
Myoglobin binds to O2 so strongly (as seen in figure 5.8) that little O2 would be released in tisuse in spite of a drop in pressure. Hence, the O2 affinity at low partial pressures is too high.
5.2.2.1 Allostery
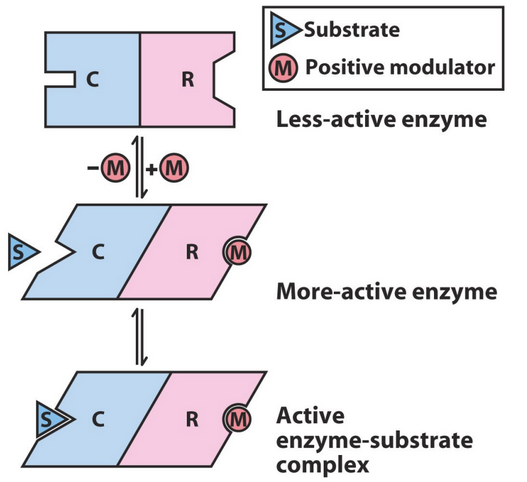
Figure 5.9: Allosteric Effect in Action
An allosteric effect (see figure 5.9) is a molecular binding event that alters the conformation and / or the dynamics of a macromolecule at a distant location.
Allosteric effects are ubiquitous in Biology. They are necessary to for coorperative behavior in hemoglobin and many other systems.
Some other examples of allostery include enzyme activation via effector binding at a distant location, hence resulting in structural changes that promote substrate binding and catalytic activity.
5.2.3 T / R state transitions
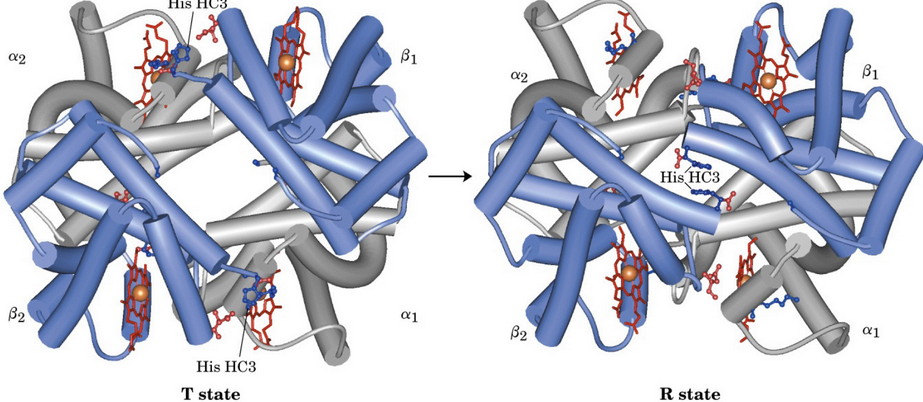
Figure 5.10: T and R States of Hemoglobin
The T state (i.e., “tense” state) of hemoglobin has a low affinity for O2 and is favored by a lack of bound O2.
The R state (i.e., “relaxed” state) of hemoglobin has a high affintiy for O2 and is favored by O2 binding.
The transition between the T and the R state of hemoglobin is mediated by the chemistry of the heme complex. When a dative covalent bond is formed between O2 and the iron cation (i.e., a Fe2+), the geometry of the heme group transitions to a more octahedral configuration that translates across the entire hemoglobin tetramer.
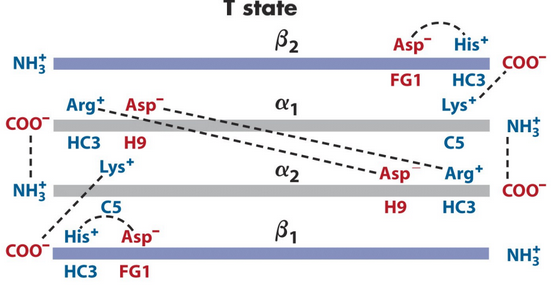
Figure 5.11: Closeup of the T State
A number of ion pairs stabilize the T state of hemoglobin in figure 5.11. When O2 binding induces the R state of hemoglobin, the \(\alpha_1\beta_1\) and the \(\alpha_2\beta_2\) subunits slide past each other and rotate, hence causing some of the T state ion pairs to be broken (notably, the HC3 carboxy-terminus with Lys C5) and allowing some new ones to be formed.
5.2.4 2,3-Biphosphoglycerate (i.e., BPG)
BPG is a highly charged molecule that selectively binds to the T state of hemoglobin (hence stabilizing the deoxy configuration of hemoglobin as well).
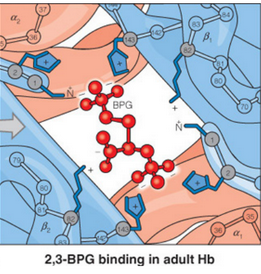
Figure 5.12: BPG Binding to the Center of the T State of Hemoglobin
As shown in figure 5.12, BPG binds to a highly positively charged central region in hemoglobin (which is only accessible in the T state). Hence, this lowers the Hb-O2 affinity and serves as part of humans’ altitude adjustment apparatuses.