9.4 Polysaccharides
Polysaccharides play roles in storage, structure, and cellular recognition.
A homopolymer or a homoglycan is a polysaccharide that only contains one kind of monosaccharide. This is in contrast to a heteropolymer or a heteroglycan: a polysaccharide that has different kinds of monosaccharides.
Starch and glycogen are homopolymers that serve storage purposes. Chitin and cellulose are structural molecules; cell surface polysaccharides are generally used as cell recognition molecules.
9.4.1 Amylose and amylopectin
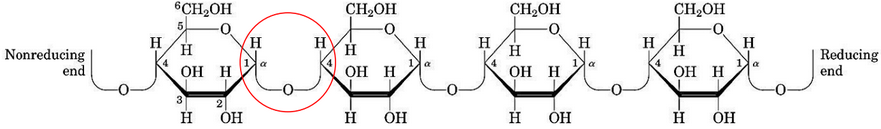
Figure 9.22: Structure of Amylose
Amylose is a long, unbranched chain of D-glucose residues and is connected by \(\alpha\)(1, 4) linkages (circled in red in figure 9.22).
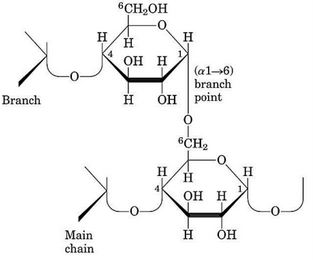
Figure 9.23: Structure of Amylopectin
Amylopectin is the branched version of amylose (see figure 9.23) and is attached via \(\alpha\)(1, 6) (i.e., the branched component) and \(\alpha\)(1, 4) glycosidic bonds (i.e., the linear component).
Amylopectin is branched once every 25 residues; the side chains of amylopectin contain 15 to 25 glucose residues.
9.4.2 Plant starch and animal glycogen
Figure 9.24: Starch in Storage Granules
Starch (see figure 9.24) is a homopolymer of glucose that is present in plant cells as a mixture of amylose and amylopectin. It is stored in granules that are 3 - 100 \(\mu m\) in diameter.
Raw starch granules resist enzymatic hydrolysis, but cooking causes them to absorb water and swell. Swollen starch is also a substrate for two different glycosidases.
Figure 9.25: Glycogen Stores in a Hepatocyte
Glycogen (see figure 9.25) is a branched homopolymer of glucose that can contain up to 50000 glucose residues.
In mammals, glycogen accounts for up to 10% of liver mass and 2% of muscle mass.
9.4.3 Cellulose
Cellulose is a water-insoluble substance that is naturally found in the cell walls of plants (e.g., stalks, stems, and trunks).
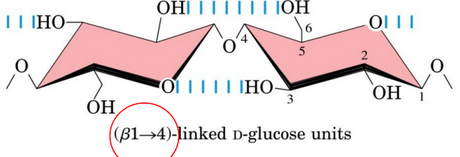
Figure 9.26: Structure of Cellulose
Cellulose molecules (seen in figure 9.26) are linear and unbranched, linked via \(\beta\)(1, 4) linkages, and can consist of up to 10000 to 15000 D-glucose units.
Note that glucose in cellulose exist in the \(\beta\) conformation; sugars in amylose and amylopectin exist in the \(\alpha\) conformation.
Furthermore, while amylose is present in a helical conformation, cellulose can exist in a fully extended conformation.
9.4.4 Chitin
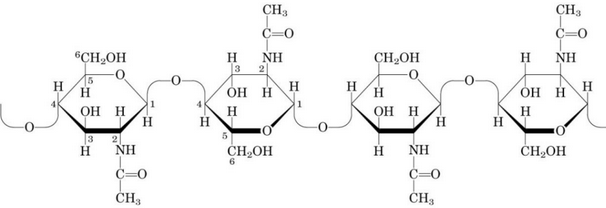
Figure 9.27: Structure of Chitin
Chitin is a homopolymer of N-acetylglucosamine residues. Like cellulose, it is also linked via \(\beta\)(1, 4) linkages and forms extended fibers similar to those of cellulose.
It is the principal of the hard exoskeletons of arthropods and is the second most prominent polysaccharide in nature (next to cellulose).
Removing an acetyl group from chitin yields a new polymer: chitosan (see figure 9.28):
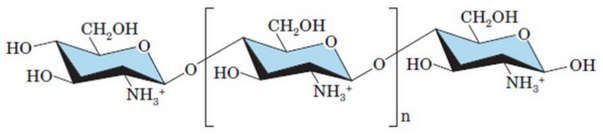
Figure 9.28: Structure of Cellulose
Due to the NH3+ groups in chitosan, blood coagulation can be accelerated by attracting negatively-charged platelets to form a clot.
When platelets aggregate, they activate coagulation molecules (e.g., thrombin, serotonin, ADP, and TxA2) that accelerate the aggregation of fibrin and red blood cells to form a coherent blood clot to plug injured blood vessels.
9.4.5 Bacterial cell wall
All bacteria have a strong, protective peptide-polysaccharide layer called the peptidoglycan.
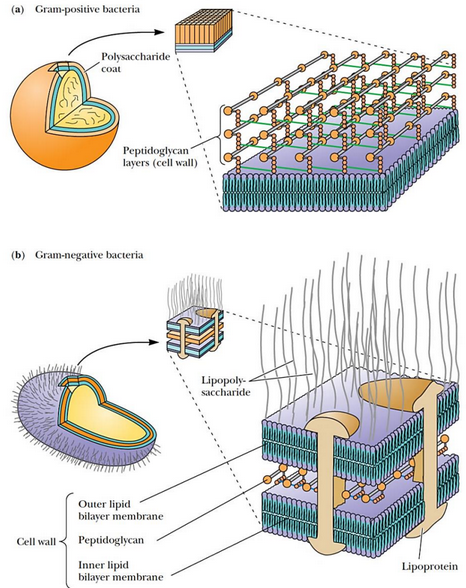
Figure 9.29: Thickness of Cell Wall in Gram Postiive and Gram Negative Bacteria
In gram positive bacterium, the cell wall is about 25 mm thick; the cell wall is about 2 to 3 nm thick in gram negative bacterium (see figure 9.29 above).
Also note that the peptidoglycan is sandwiched between an inner and an outer layer of lipid bilayers.
9.4.5.1 Constituents of peptidoglycan
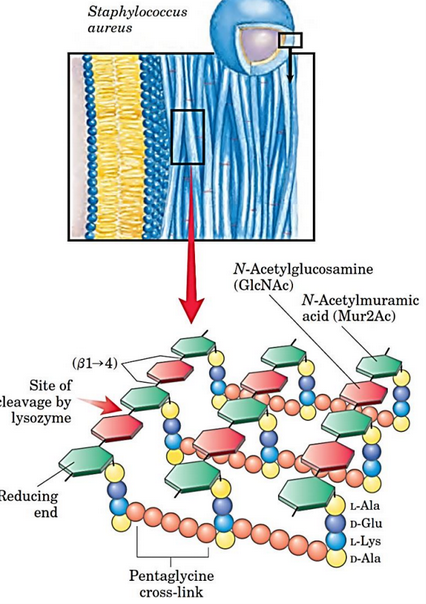
Figure 9.30: Peptidoglycan Composition
Peptidoglycan is a heteropolymer of alternating N-acetylglucosamine (i.e., GlcNAc) and N-acetylmuramic acid (i.e., Mur2Ac) residues. It is also linked via \(\beta\)(1, 4) linkages (and hence, is linear).
The peptide cross links in figure 9.30 creates a strong sheath that envelops the bacterium and prevents cellular swelling and lysis (from osmosis).
The enzyme lysozyme kills bacteria by hydrolyzing the \(\beta\)(1, 4) glycosidic bond in peptidoglycan. Peptidoglycan is also structurally similar to chitin, albeit it is joined to a tetra peptide (usually L-Ala, D-Glu, L-Lys, and D-Ala); the L-Lys is linked to the \(\gamma\)-COOH of the D-Glu residue:
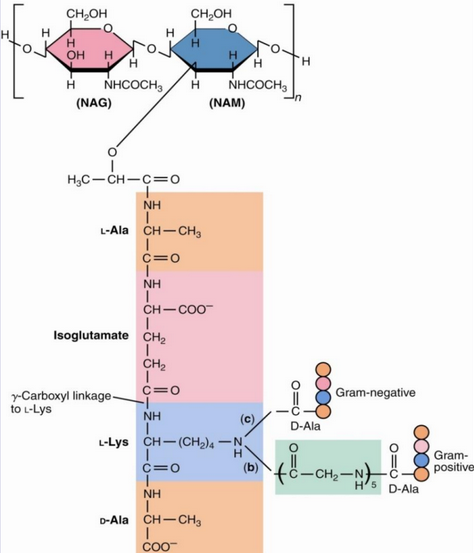
Figure 9.31: Tetra Peptides in Peptidoglycan
The tetra peptide is linke to the NAM (i.e., N-acetylglucosamine) units in figure 9.31 via its D-lactate moiety. The \(\epsilon\)-amino group of lysine in this peptide is also linked to the COOH group of D-alanine of an adjacent tetra peptide.
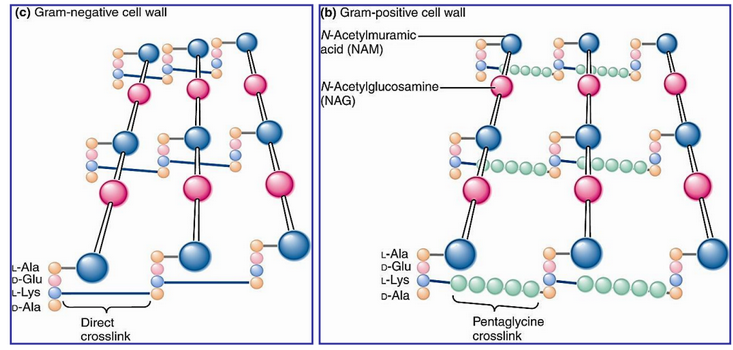
Figure 9.32: Crosslinks in Gram Positive and Gram Negative Bacteria
Also note that the lysine in gram negative cell walls forms a direct amide bond with the carboxyl group of D-alanine.
In gram positive cells, a pentaglycine chain (see figure 9.32) bridges the lysine and the D-Ala carboxyl group.
9.4.6 Glycoconjugates
Glycoconjugates are oligosaccharides that have been covalently linked to lipids and / or proteins. These glycoconjugates are present on the cell surface and are responsible for cell-to-cell recognition.
Some examples of glycoconjugates include:
- Proteoglycans
- Glycoproteins
- Glycolipids
9.4.7 Glycosaminoglycans
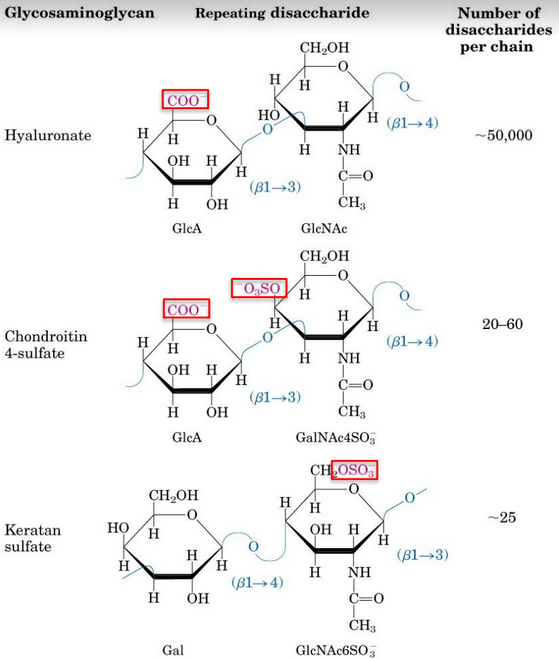
Figure 9.33: Disaccharides in Glycosaminoglycans
Glycosaminoglycans is a polysaccharide is involved in a variety of extracellular (and sometimes, intracellular) functions.
Glycosaminoglycans are also fundamental constituents of proteoglycans; glycosaminoglycans are represented by linear chains. One of the monosaccharide units is an amino sugar; the other monosaccharide units contain at least one negatively charged sulfate or carboxylate group.
9.4.8 Proteoglycans
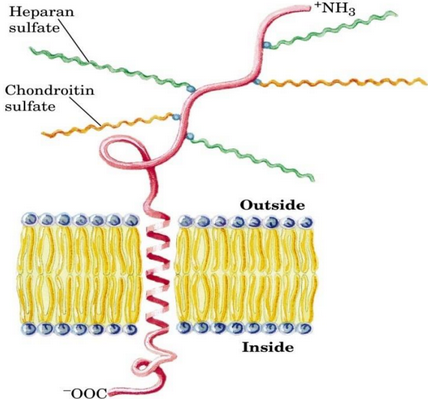
Figure 9.34: General Schematic of a Proteoglycan
These are macromolecules that have been covalently joined to a membrane protein (see figure 9.34).
A trisaccharide linker connects a glycosaminoglycan residue to a serine residue in the core protein.
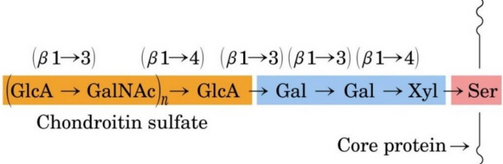
Figure 9.35: Closeup of a Proteoglycan Structure
The xylose residue at the reducing end of the linker is then joined to its anomeric carbon to the hydroxyl group of the serine residue in figure figure 9.35.
9.4.9 Glycoproteins
These are carbohydrate-protein conjugates; the carbohydrate moieties in glycoproteins are smaller than the glycosaminoglycan moiety. There is also a glycosidic linkage between the two moieties (observe figure 9.36):
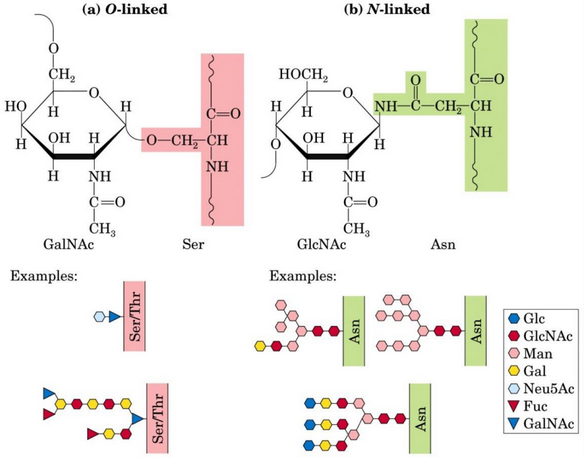
Figure 9.36: Glycoproteins Linked by Glycosidic Bonds
When an O-glycosidic bond is present, the anomeric carbon atom interacts with the OH group of serine or threonine. Otherwise, if a N-glycosidic bond is present, the anomeric carbon atom interacts with the amide nitrogen of the asparagine residue.
9.4.10 Glycolipids and lipopolysaccharides
Glycolipids are membrane lipids with hydrophilic oligosaccharides as their head groups.
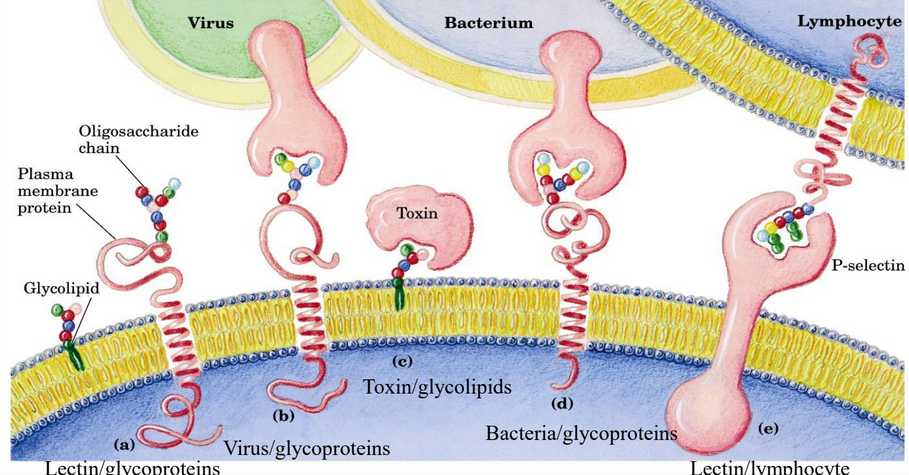
Figure 9.37: Oligosaccharides in Recognition and Adhesion Processes
Nonetheless, glycolipids also serve as specific recognition sites for carbohydrate binding proteins (see figure 9.37). Different oligosaccharides are present in the four blood types in humans.
Blood types in humans also depend on the type of ganglioside present on the surface of the red blood cell.