3.2 Secondary Structures
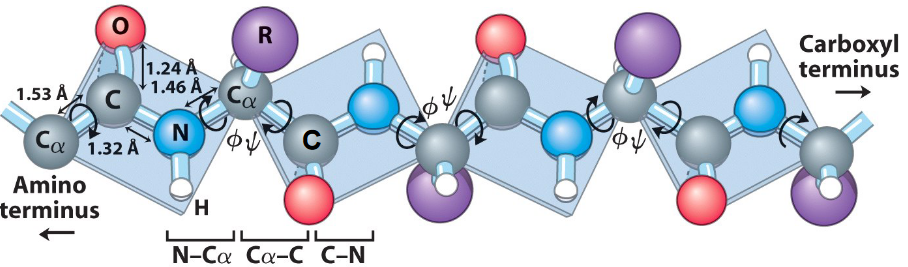
Figure 3.6: Amide and Peptide Bond Planes
All amide and peptide bond planes are joined by the tetrahedral bonds of the \(\alpha\) carbons (see figure 3.6). There are also two rotational parameters:
- \(\psi\) represents the angle about the C\(\alpha\)-C bond
- \(\phi\) represents the angle about the C\(\alpha\)-N bond
Hence, the entire path of the peptide will be known if any only if all \(\psi\) and \(\phi\) angles are known. Some values of \(\psi\) and \(\phi\) angles are more common than others.
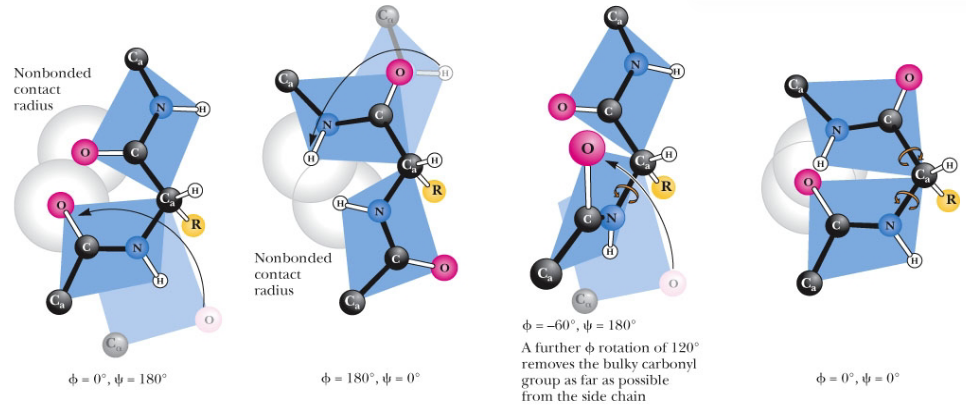
Figure 3.7: Forbidden Values of Psi and Phi
However, do note that certain values of \(\psi\) and \(\phi\) angles are not permitted due to steric hindrance (see figure 3.7)!
3.2.1 Ways of folding the polypeptide chain
Pauling et. al. suggested several principles that all polypeptide structures must obey:
The bond lengths and the bond angles should be distorted as little as possible from those found through X-ray diffraction studies of amino acids and peptides.
No two atoms should approach one another more closely than their van der Waals radii.
The amide group must remain planar and in the trans configuration. Hence, rotations are only possible about the two bonds adjacent to the \(\alpha\)-carbon in each amino acid residue.
Noncovalent interactions are necessary to stabilize regular folding. For instance, hydrogen bonding between amide protons and carbonyl oxygens help stabilize the shape of \(\alpha\)-helices.
3.2.2 Types of secondary structures
All secondary structures are local structures that are stabilized by hydrogen bonding!
The secondary structures focused on in BS1005 / CM1051 are:
- \(\alpha\)-helices
- \(\beta\)-sheets
- Tight turns
- \(\beta\)-bulges
3.2.2.1 \(\alpha\)-helices
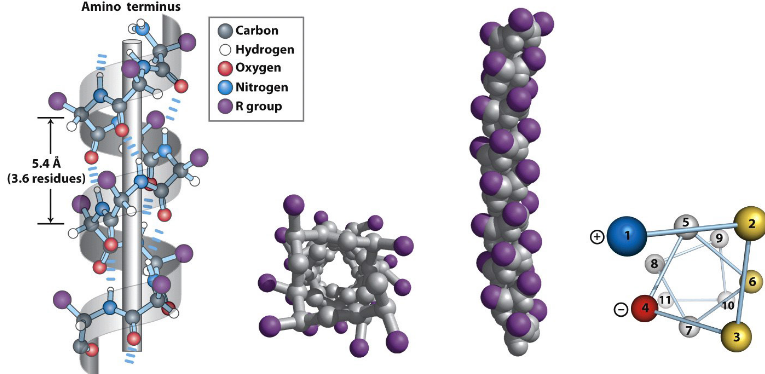
Figure 3.8: An Amphipathic Alpha Helix
The alpha helix (see figure 3.8) was first proposed by Linus Pauling and Robert Corey in 1951 (and identified in keratin by Max Perutz). It is an integral component of proteins and is stabilized by hydrogen bonds.
An \(\alpha\)-helix has 3.6 amino residues per turn and a rise of 1.5 Angstroms per residue. Hence, the rise per pitch of the \(\alpha\)-helix is 5.4 Angstroms.
The backbone loop of an \(\alpha\)-helix is made up of 13 atoms. The hydrogen bonds in an \(\alpha\)-helix are also almost parallel to the helix’s axis.
The torsion angles are around \(\psi = -45^\omicron\) and \(\phi = -60^\omicron\) for this secondary structure.
3.2.2.1.1 3.613 and 310 helices
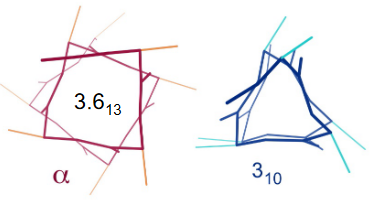
Figure 3.9: Other Helices to Note
In a 3.613 helix, there is a loop of 13 amino acids; each turn of the helix has 3.6 amino acids. Hydrogen bonds in the helix tend to be linear, hence granting the helix stability.
In contrast, the 310 helix has exactly three residues per turn and a 10-atom hydrogen-bonded loop.
3.2.2.1.2 Net dipoles in \(\alpha\)-helices
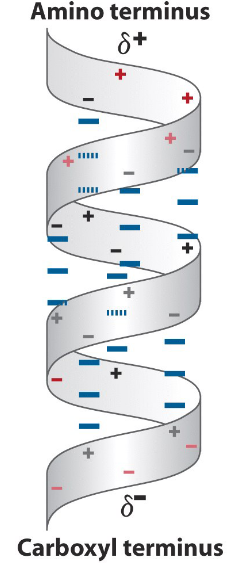
Figure 3.10: Net Dipoles in Alpha Helices
The arrangement of N-H groups pointing towards the N-terminus and the C=O groups pointing towards the C-terminus creates a significant net dipole moment for the helix (as observed in figure 3.10).
3.2.2.2 \(\beta\)-pleated sheets
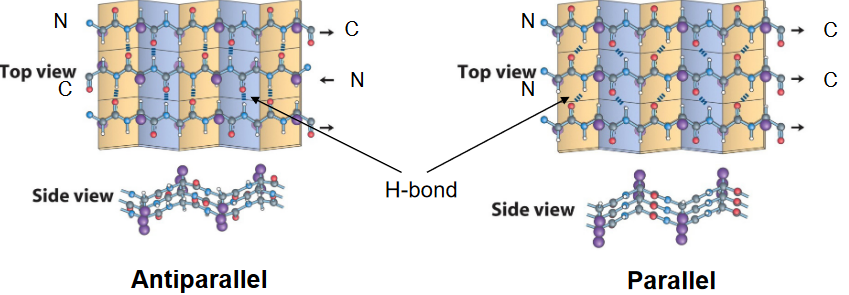
Figure 3.11: Parallel and Antiparallel Beta Sheets
This is also a structure that was first postualted by Pauling and Corey in 1951. Nonetheless, the \(\beta\)-pleated sheet is made up of \(\beta\)-strands and may be parallel or anti-parallel to one another (see figure 3.11).
Each \(\beta\)-sheet is made up of two or more strands, and each residue in the strand is rotated by 180\(^\omicron\) with respect to the preceding one.
The two strands can also be arranged such that the N-terminus to the C-terminus orientations are in opposite directions (i.e., “antiparallel”) to one another. The hydrogen bonds between anti-parallel strands are linear.
Alternatively, the two strands can also be arranged such that the N-terminus to the C-terminus orientations are in the same direction (i.e., “parallel”). However, the hydrogen bonds between parallel strands are not linear!
3.2.2.3 \(\beta\)-turns
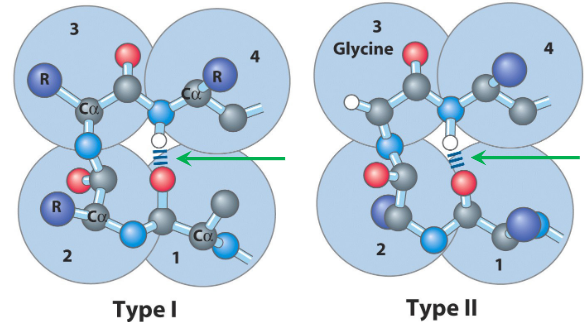
Figure 3.12: Two Types of Beta Turns
\(\beta\)-turns are what allows a peptide chain to change direction. Type I turns are the most common; type II turns (as seen in figure 3.12) have a third glycine residue!
Four residues are required to form a \(\beta\)-turn, and the oxygen of a carbonyl group of one residue is hydrogen bonded to the amide proton of a residue three residues away.
The amino acids proline and glycine are also prevalent in \(\beta\)-turns.
3.2.3 Steric constraints on \(\phi\) and \(\psi\)
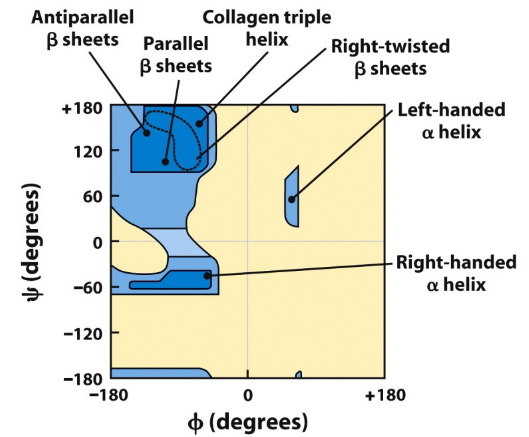
Figure 3.13: Steric Constraints on Torsion Angles phi and psi
The Ramachandran diagrams above show the allowed combinations of the torsional angles \(\psi\) and \(\phi\). Blue regions in figure 3.13 are favorable angle combinations.
However, the blue regions on figure 3.13 do not account for angles allowed for the amino acid glycine - this is because glycine is a particularly flexible amino acid (as it does not have a “R” group).
3.2.4 Circular dichroism spectroscopy
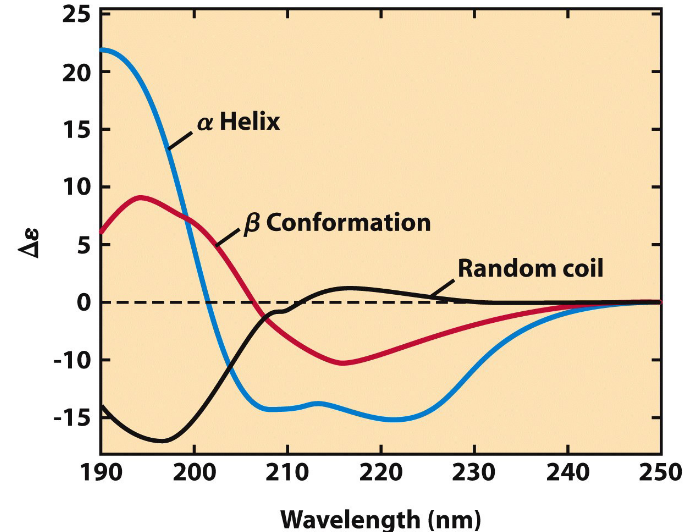
Figure 3.14: Circular Dichroism Spectroscopy
In circular dichroism spectroscopy, one measures the molar absorption difference \(\Delta \epsilon\) between left and right circularly polarized light - that is, \(\Delta \epsilon = \epsilon_L - \epsilon_R\).
This spectroscopy spectrum can also provide an approximate estimation for the fraction of general secondary structures assumed in a given protein. The spectrum of a native protein can also serve as a benchmark for the folded state, which is useful for monitoring protein denaturation or conformational changes brought about by changes in the environment.