8.5 Biological Functions of RNA
8.5.1 RNA viruses
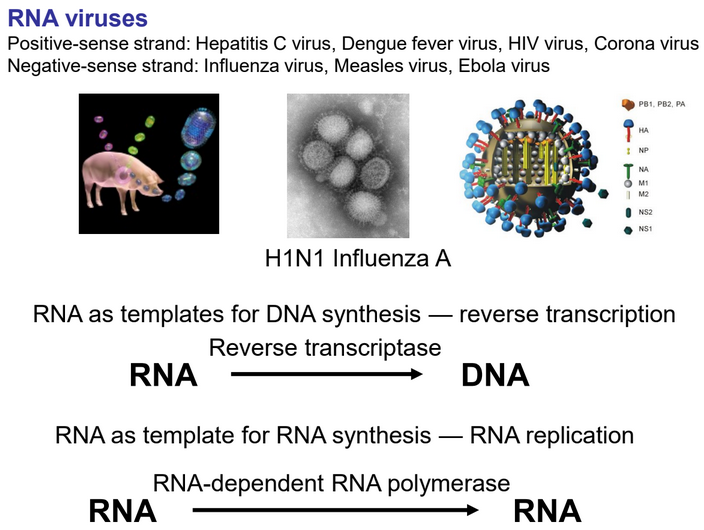
Figure 8.13: Several RNA Viruses
RNA can serve as a mode of genetic information storage in viruses (i.e., a RNA genome).
8.5.2 Protein synthesis
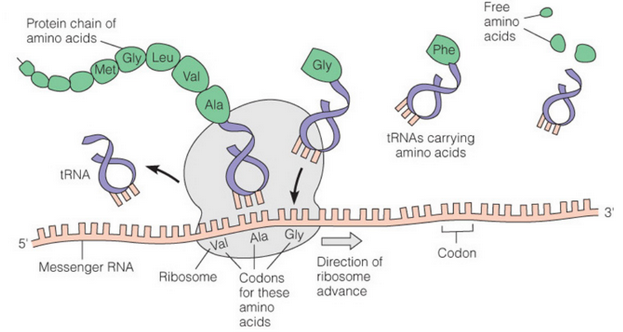
Figure 8.14: RNA Molecules in Protein Synthesis
In figure 8.14, mRNA carries coding information to the from a gene to the ribosomes (with rRNA as its major components).
The ribosome then binds to the mRNA and tRNAs then bring amino acids to the ribosomes one at a time.
Each tRNA is matched to the codon on the mRNA, and the ribosome adds the corresponding amino acid to the growing protein chain. The ribosome then migrates along the mRNA to read the genetic message and translate it into a protein.
8.5.3 Ribozymes
In an experiment by Thomas Cech et al. in 1981, Cech and his colleagues discovered that during the synthesis of 26S rRNA in Tetrahymena (i.e., a ciliated protozoan), a 0.4 kb intron (i.e., non-coding RNA) was removed.
To determine whether or not enzymes were responsible for this loss, the splicing activity was carried out again with purified 26S rRNA in the presence and in the absence of a nuclear abstract. The results proved that splicing could also occur in the absence of a nuclear abstract.
Heat denaturation, chemical denaturation, and even proteinase treatment was also carried out to rule out the possibility that the purified rRNA could have been contaminated with proteins. However, splicing activity was still observed. Hence, it was determined that 26s rRNA had self-splicing activity.
Nonetheless, a ribozyme is a general term used to refer to RNA with catalytic activity (i.e., a “RNA enzyme”). Whereas some ribozymes are directed against other substrates, others have intramolecular activities that limit its catalytic action to a single cycle.
8.5.4 RNA splicing (group I intron mechanism)
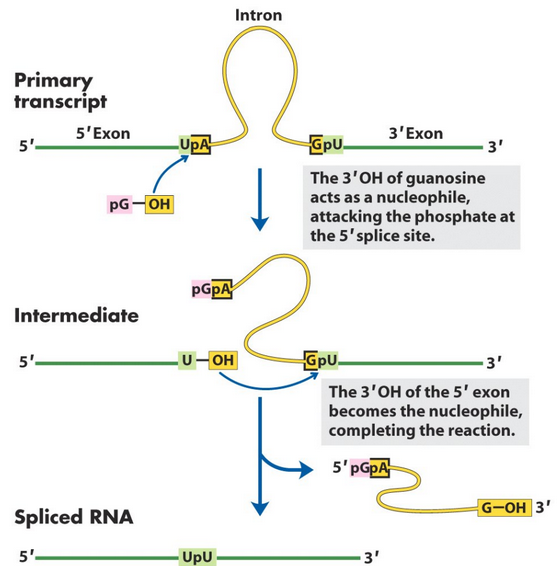
Figure 8.15: Group I Intron Splicing Mechanism
Some introns are able to splice themselves out of the mRNA that contains them via the mechanism in figure 8.15. This is also an automated way to separate the coding exon(s) from the non-coding intron(s).
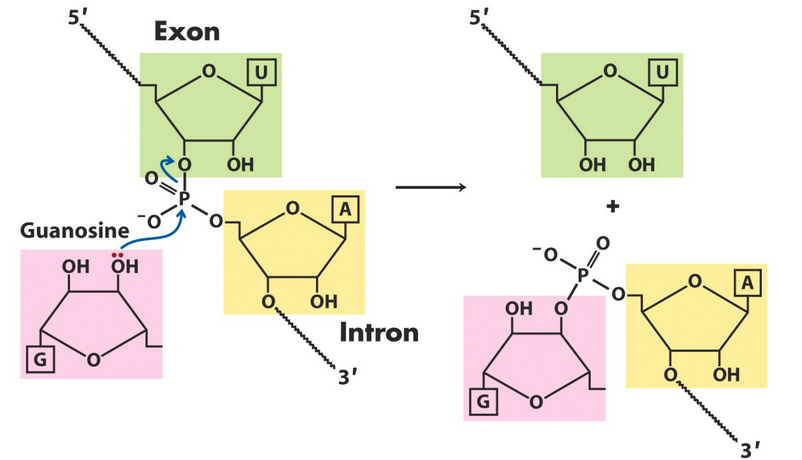
Figure 8.16: One of Two Splicing Reactions
The group I intron mechanism has two transesterification reactions, the first of which is shown in figure 8.16.
8.5.5 Ribonuclease P
The enzyme ribonuclease P (i.e., RNAse P) is a ribonucleoprotein that has a single RNA molecule bound to a protein (see figure 8.17)
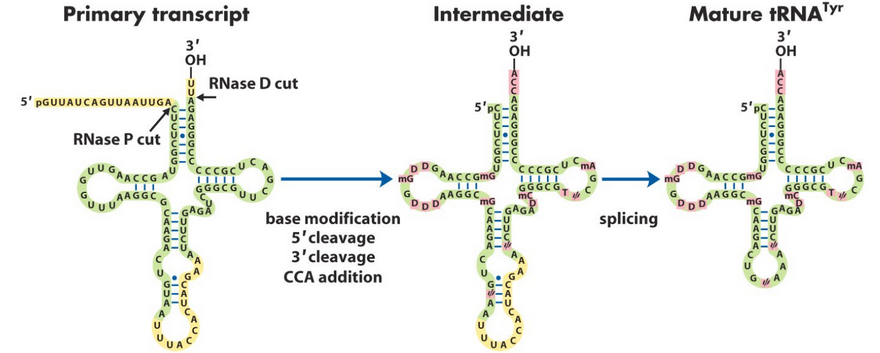
Figure 8.17: Ribonuclease P Activity
This protein has the ability to catalyze the cleavage of a pre-tRNA substrate. Funnily enough, this protein also plays a peripheral role - perhaps in stabilizing catalytic RNA.
8.5.6 Hammerhead ribozymes
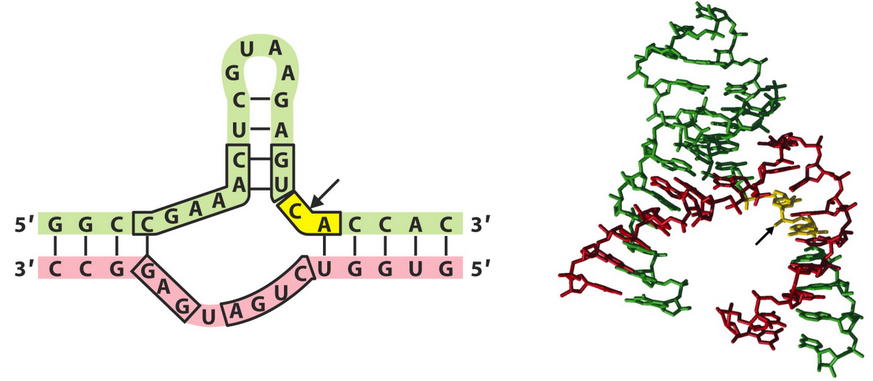
Figure 8.18: Structure of a Hammerhead Ribozyme
The hammerhead structure in figure 8.18 is formed via pairing interactions between a substrate strand (i.e., the green strand in figure 8.18) and an “assister” strand (i.e., the red strand in figure 8.18).
The hammerhead ribozyme is a metallo-catalyst: it requires a magnesium ion (i.e., Mg2+) for catalytic activity.
8.5.6.1 Catalytic mechanism for hammerhead ribozymes
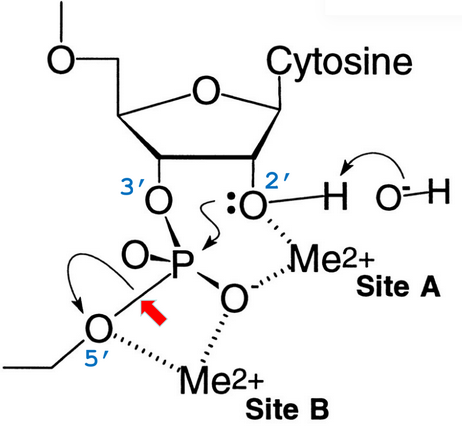
Figure 8.19: Two Metal Ion Model of Ribozymes
The divalent ion at site A of figure 8.19 coordinates the 2’ oxygen to weaken the 2’ OH bond by promoting it to the alkoxide form - 2 O’ - the superior nucleophile.
The divalent ion at site B of figure 8.19 also promotes the development of the negative charge on the leaving 5’ oxygen (which weakens the 5’ O-P bond).
The metal ion in vivo would typically be a Mg2+ ion.
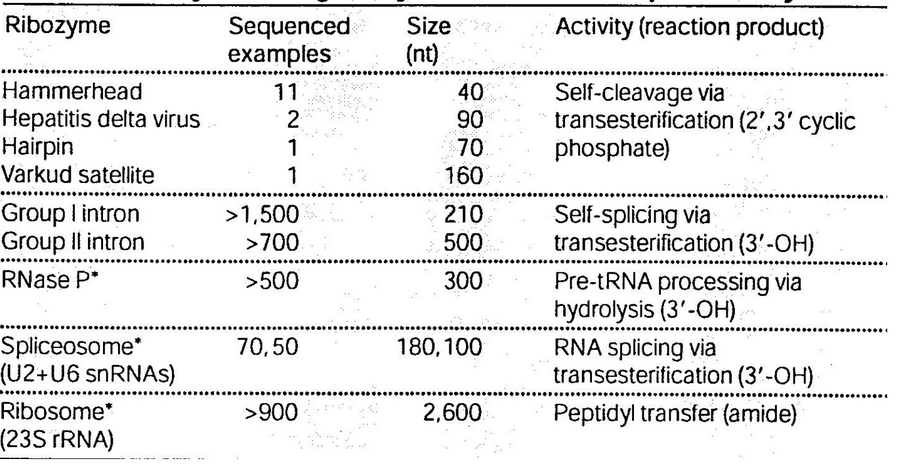
Figure 8.20: Summary of Naturally-Occuring Ribozymes and Ribonucleoproteins Enzymes
Figure 8.20 above shows the summary information of naturally occurring ribozymes and ribonucleoprotein enzymes.
8.5.7 Small non-coding RNAs
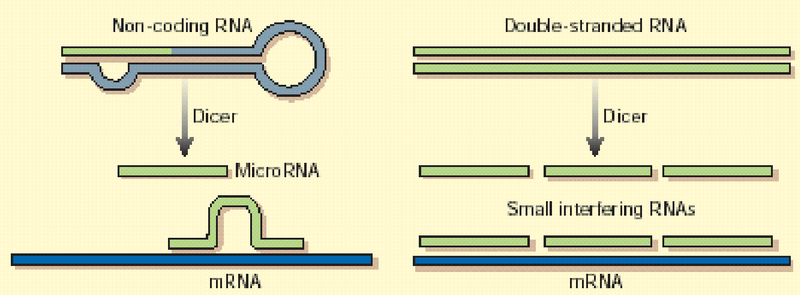
Figure 8.21: ncRNAs at Work
Only less than 1.5% of human DNA codes for protein (i.e., the exons). Strangely enough, 25% of the human genome codes for “non-coding RNAs” (i.e., ncRNAs).
There are also two important classes of small, non-coding RNAs - they are:
MicroRNA (i.e., miRNA)
These are small (about 20 nucleotides long) and are single-stranded.
Small interfering RNA (i.e., siRNA)
These are also about 20 nucleotides long, but are double-stranded.
Both miRNAs and siRNAs are invovled in gene regulation by binding to mRNAs (see figure 8.21)!