Capítulo 4 Fatores de Suscetibilidade às Doenças
Muitos são os fatores que podem modificar a suscetibilidade de um animal às doenças. Nós podemos classificá-los em dois grandes grupos. Os fatores abióticos são os componentes do ambiente supra-biológicos e exercem influência sobre os organismos biológicos, determinando os limites de sua viabilidade ecológica, e afetando seu bem-estar e saúde de várias formas. Já os fatores bióticos são toda e qualquer influência exercida sobre a saúde de um animal pelos outros organismos vivos que com ele podem entrar em contato, e interagir, durante o curso de vida do animal. No curso do presente capítulo eu discutirei alguns destes fatores, selecionados por sua importância ou abrangência de influência.
4.1 Fatores abióticos
A evolução biológica da Terra acompanhou a geológica; foram e são, em última instância, os fatores abióticos que determinam as condições de viabilidade biológica em qualquer ecossistema planetário. Em nosso caso em particular, a partir da formação do planeta Terra há 4,5 bilhões de anos atrás, foi a mudança gradativa de temperatura, radiação, formação e gradativa mudança de constituição atmosférica, formação e mudança de constituição da crosta, que moldaram a evolução biológica no planeta até os tempos modernos. Durante todo este tempo geológico, tais mudanças de fisionomia dos ambientes geraram fatores de seleção natural completamente diferentes em local e tempo, acarretando tanto grandes extinções biológicas como também grandes explosões de diversidade de vida posteriores às extinções. Entretanto, a análise feita por palentologistas do curso do tempo destas extinções demonstra que a recuperação pode levar muito tempo, já que demora para que genomas bem sucedidos consigam estabelecer-se e dominar o pool resultante no novo ambiente. Estabilidade ambiental e diversidade biológica andam juntos; alterações ecológicas abruptas ou radicais acarretam mudanças na biota. As doenças emergentes atuais são um exemplo atual das consequências da alteração antropocêntrica do ambiente.
4.1.1 Água
A água é uma molécula fundamental para os processos biológicos em nosso planeta. A Terra tem 75% da superfície coberta de água, e é comumente chamada pelos humanos de “Planeta Água”. Esta percepção de que a água está e estará sempre prontamente disponível, que tal composto é inesgotável, não poderia ser mais equivocada no mundo atual de 7 bilhões e 700 milhões de pessoas. Observe a Figura 4.1: nela você pode notar que de toda o volume de água que existe no planeta, grande parte é salgada (nos oceanos em geral), ou inacessível (em glaciares e aquíferos em geral). Assim, a água que temos efetivamente disponível para beber, viver, produzir, excretar, plantar, colher, criar e abater é apenas parcela milesimal do total existente em nosso planeta e esta quantidade pequena tem uma distribuição muito desigual através dos continentes (Thomas 1994).
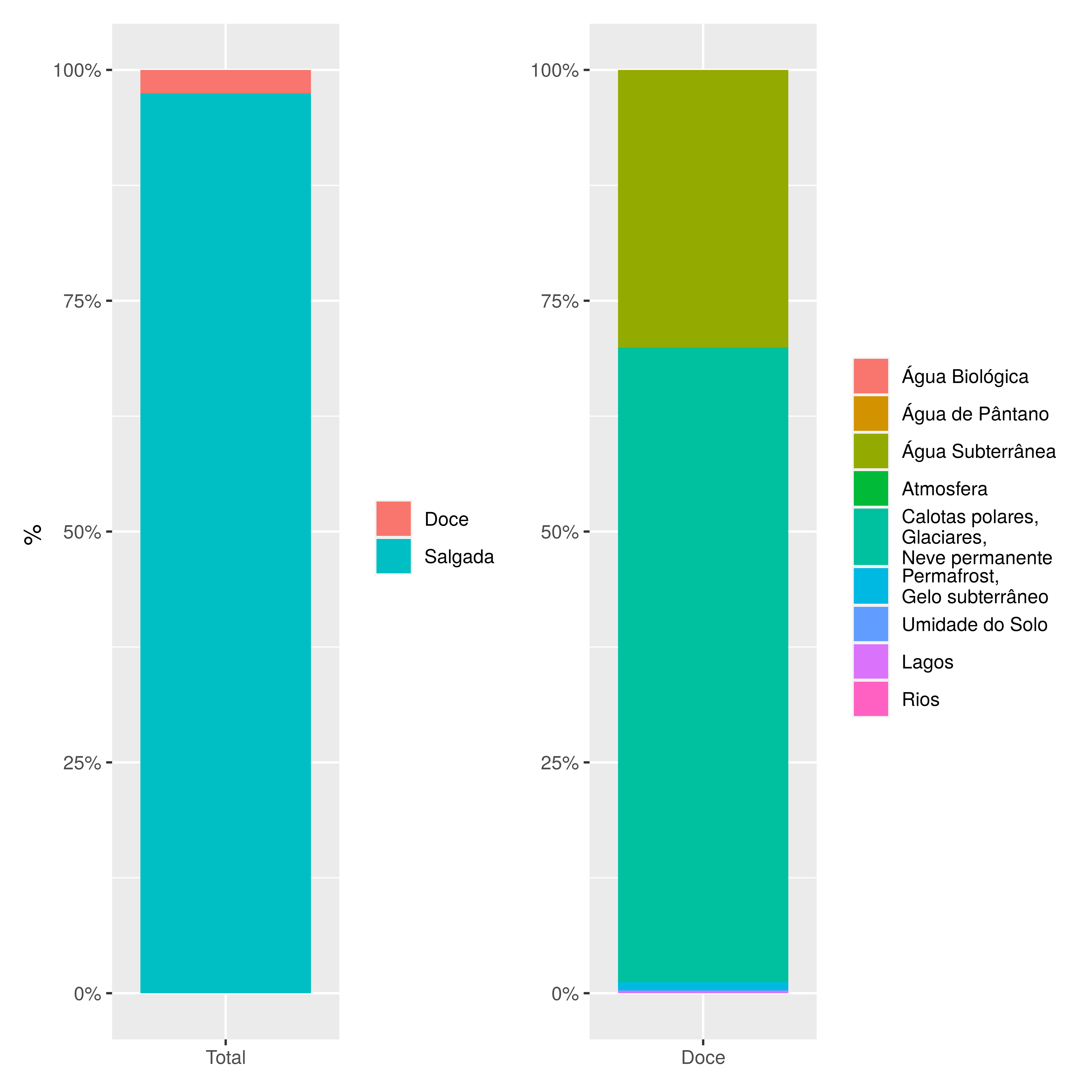
Figura 4.1: Particionamento da água no planeta Terra. A) O volume de água doce no planeta inteiro equivale a apenas 2,5% da água total. B) O volume de água doce disponível para uso humano, oriunda na maioria de Lagos e Rios, equivale a 0,007% (ou 93.120 Km3) da água total.
E a água é fundamental no que tange à prevenção, diagnóstico e controle e prevenção de doenças. Sem água de qualidade para limpeza, bebida e realização de testes diagnósticos todo o controle de doenças de um plantel de uma residência ou propriedade fica ameaçado. Nesta seção, é de nosso interesse porquê a água é importante para os organismos biológicos em si. Isto porquê ela possui duas propriedades fundamentais: solvente universal e tampão de temperatura.
4.1.1.1 Solvente universal
A água é uma molécula que, devido a sua estrutura tetraédrica (vide Figura 4.2) comporta-se como um dipolo. Graças a esta característica as moléculas água são capazes de ligar-se através de pontes de hidrogênio tanto entre si, como entre biomoléculas e íons aceptores ou doadores de elétrons, estabilizando as biomoléculas e solubilizando os íons. Portanto, a molécula da água pode reagir com praticamente qualquer molécula, desde que ela apresente carga iônica. Moléculas apolares, como lipídios em geral, são incapazes de reagir com as moléculas de água. Como resultado dos processos milenares de evaporação, condensação e precipitação da água na atmosfera, com consequente aglomeração em coleções e cursos d´água, ocorre a lixiviação e erosão das rochas de nosso planeta. Tais processos são resultado das reações químicas entre a água e tais rochas, solubilizando e disponibilizando os vários minerais componentes para uso biológico (Murray and Harper 1996).
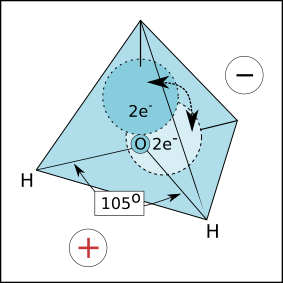
Figura 4.2: Molécula da água. Devido ao agrupamento de elétrons no vértice do tetraedro mais distante dos dois hidrogênios a molécula da água pode reagir com íons negativos e positivos, permitindo a ela tornar-se Solvente Universal.
Dentro dos organismos vivos a água cumpre a mesma função, solubilização de minerais e outras moléculas, além da condução de gases como O2 e CO2. É por isso que a grande maioria dos organismos vivos é composto por 70% a 76% de água: para que bioproteínas, enzimas e íons possam circular e reagir entre si (Culebras et al. 1977). Lembre-se que a água biológica é compartimentalizada em água intra-celular, intersticial e intra-vascular, e que o fluxo de água entre esses compartimentos pode sofrer modulações tanto da ‘membrana’ capilar como da membrana celular. Em última instância a manutenção do equilíbrio hídrico dos animais é essencial para a viabilidade biológica.
É a dissolução de CO2 e O2 na água que permite a vida aeróbica em coleções d´água e permite a condução destes compostos de e para a célula em organismos pluricelulares através de sistemas circulatórios. Todavia, a capacidade da solução de água reter compostos gasosos é diretamente dependente de temperatura da solução. Quanto mais quente a água, menos oxigênio e gás carbônico ela é capaz de reter. Como o gás O2 dissolve-se na água com mais dificuldade do que o CO2, o primeiro é muito mais afetado. Assim um mesmo volume de água a uma temperatura maior sustenta uma Demanda Biológica de Oxigênio (DBO) menor; menos vida haverá em tal volume, portanto. Este é um dos mais sérios efeitos do aquecimento global sobre as coleções d´água do planeta.
4.1.1.2 Tampão de Temperatura
A segunda característica da molécula da água fundamental para a vida em nosso planeta é que ela age, no planeta e nos organismos biológicos, como um Tampão de Temperatura. Dependendo de seu estado (sólido, líquido ou gasoso) a água é capaz de absorver quantidades diferentes de temperatura, conforme demonstrado na Figura 4.3. Nela observe que, para aquecermos um mol de água congelada para o estado líquido é necessário fornecer 334 Joules de energia calórica. A partir daí e até chegar à temperatura de Ebulição (100oC nas CNTP) este mol de água precisará absorver mais de 4J para cada grau Celsius de elevação de temperatura do mol de água, 420J no total. Já para aquecermos este mol de água para que ele saia do estado líquido e passe para o gasoso (evapore) ele precisará absorver incríveis 2265J de energia (Murray and Harper 1996).
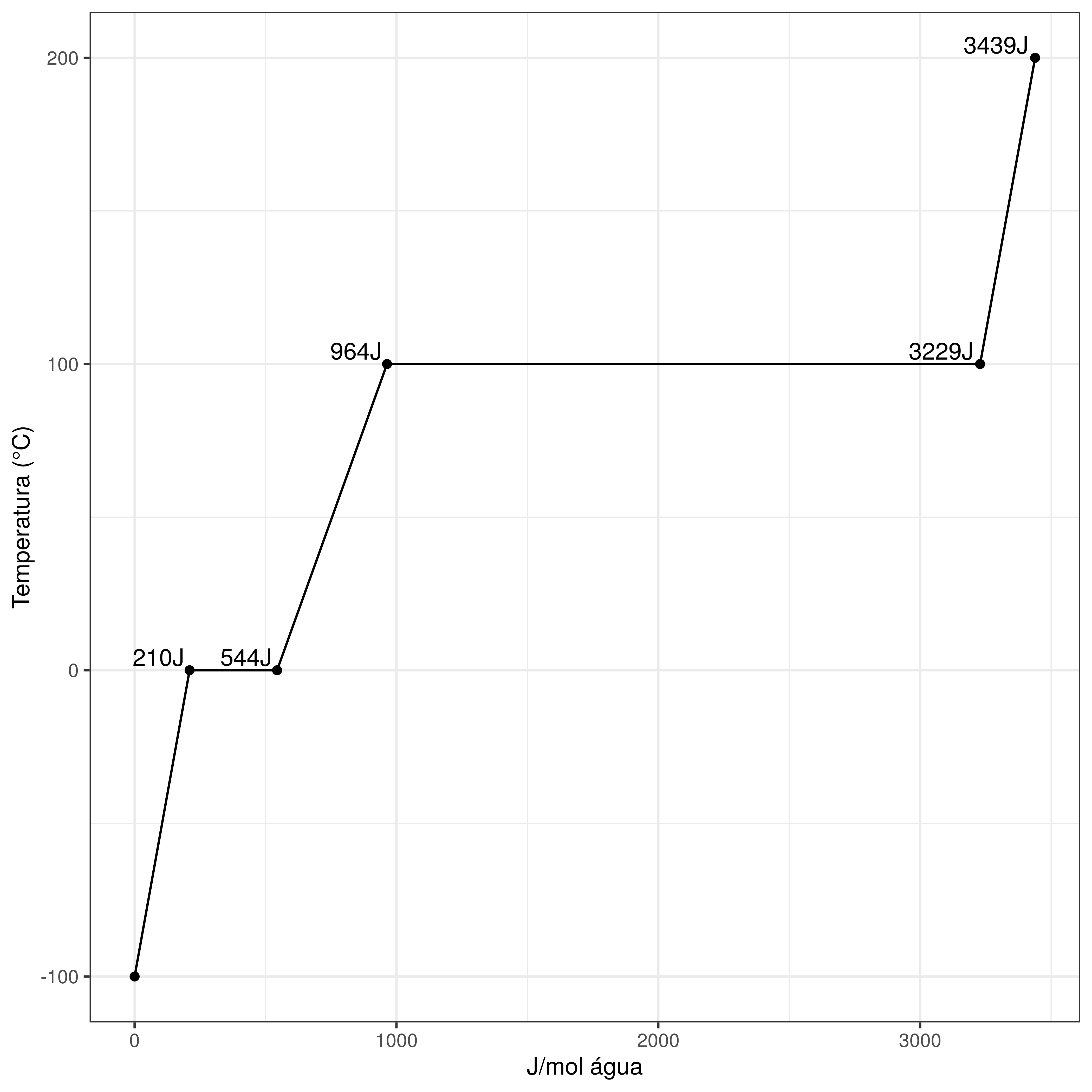
Figura 4.3: Gráfico demonstrando as variações na energia necessária para que um mol de água seja aquecido de -100oC (francamente congelada) até 200oC.
Esta capacidade de reter calor molda as flutuações de temperatura nos diversos ambientes no planeta: quanto menos água no ambiente, maiores as flutuações de temperatura diárias e no curso das estações do ano. Ambientes tropicais úmidos apresentam temperaturas ambientais relativamente altas às regiões temperadas, mas as flutuações de temperatura tem amplitude maior; assim, digamos nos remanescentes de florestas tropicais úmidas planetárias as temperaturas podem atingir os 40oC durante o dia, mas raramente caem abaixo dos 20-25oC durante a noite. Já em um deserto, você atinge os mesmos 40oC durante o dia, mas à noite a temperatura cai até os -40oC. Assim, o processo de aquecimento global agora em curso está tamponando, através do derretimento das calotas polares, Antártida, Groenlândia, Permafrost, uma quantidade imensa de energia que está sendo produzida pelos processos tecnológicos humanos.
Já nos organismos biológicos, o controle de temperatura interna celular é fundamental para que seu metabolismo possa acontecer de forma adequada (Seebacher 2009). Boa parte deste metabolismo depende de processos enzimáticos. As taxas de transformação (ou conversão) de substrato em produto molecular das enzimas (peptídeos endógenos capazes de favorecer ou propiciar clivagens, cisões e transposições moleculares) são altamente dependentes da temperatura ambiente. Toda enzima possui uma faixa ótima de temperatura de funcionamento, dentro da qual sua taxa de transformação é máxima. Abaixo da faixa ótima, a taxa de conversão cai abruptamente, até que cesse (devido à extrema baixa temperatura). Logo acima da faixa ótima de temperatura há uma estabilização da conversão, até que, se a temperatura continuar a subir, a enzima entra em processo de desnaturação, primeiro de forma reversível, mas a partir de uma determinada grau de hipertemia de forma irreversível. Sendo assim, o controle de temperatura celular dos organismos vivos é fundamental para sua viabilidade ecológica, e vários sistemas orgânicos são ajustados para tentar controlar a temperatura corporal, no caso de um vertebrado (Clarke and Fraser 2004). Sudorese, evaporação salivar, aumento de vascularização das extremidades, abanamento, busca por hidratar-se ou molhar-se, todos são mecanismos utilizados pelos vertebrados para controle de temperatura. Note que no processo de evolução dos vertebrados uma maior independência da água foi um fator de seleção relevante, sendo a evolução da alça de Henle o maior salto seletivo dos últimos 100 milhões de anos. A partir da alça de Henle tanto aves como mamíferos puderam economizar água em seus processos orgânicos (com clara vantagem dos mamíferos, já que apenas uma parcela de 30% dos néfrons das aves tem alça de Henle), através da concentração da urina, economizando muita água para uso no metabolismo, e não na excreção (Keogh, Kilroy, and Bhattacharjee 2021).
4.1.1.3 Disponibilidade
Estas propriedades da água são tão importantes que que a desidratação é anormalidade orgânica que prontamente suscita respostas que levem o organismo a ingerir ou economizar água (Silanikove 2000; Fuller et al. 2020). Se grave é a desidratação, esta pode levar o organismo à morte em curto espaço de tempo; reações químicas se tornam mais difíceis, a viscosidade do meio intracelular aumenta e dificulta o trânsito iônico, a excreção de restos metabólicos fica mais difícil, a geração de energia é prejudicada pelas flutuações de temperatura. Note que tais processos ocorrem dentro das células, mas em um organismo multicelular com sistemas orgânicos especializados, são tais sistemas (excretório, circulatório, pex.) que são afetados, comprometendo o organismo como um todo.
Esta dependência dos organismos da água é um dos maiores fatores de seleção natural em nosso planeta, já que nele a distribuição e a constituição da água é desigual. Existem ambientes com água congelada, líquida, ou em vapor. Há ambientes com água quase sem outros constituintes minerais, em outros a água contém grandes quantidades de íons como Cl-, Mg2+, Ca2+, Fe2+, Na+, entre todos os outros íons minerais que existem. Há ambientes com águas com quase nenhuma vida microscópica ou contaminação química, outros com intensa contaminação biológica e constituintes que limitam os organismos que nela podem viver ou ingeri-la. Toda esta variação gera diversidade biológica, por exigir dos seres vivos adequação genotípica e portanto fenotípica para sobreviver em cada um destes ambientes.
O primeiro aspecto a ser levado em consideração é a disponibilidade da água nos ambientes, que pode variar de abundante e homogeneamente distribuída em ambientes úmidos (sejam eles tropicais, subtropicais ou temperados), até escassa e disponível apenas em locais ou estações específicas (ambientes desérticos ou com sazonalidade bem demarcada, como as savanas, caatingas e cerrados). No primeiro caso os vertebrados em geral não tem que disputar o acesso a água com outros animais e isto permite uma distribuição dispersa dos animais. Ambientes mais úmidos tendem a melhorar a sobrevivência de formas infectantes de parasitos, já que a dessecação é fator relevante para a mortalidade de tais formas. Coleções de água de lugares úmidos também tendem a acumular menos minerais em sua constituição, porquê a taxa de renovação local da água é maior.
Em ambientes secos ou marcadamente sazonais o acesso fácil à água é limitado à estação chuvosa, onde há abundância relativa de água e nesta estação é que concentram-se as estações reprodutivas animais, o que aumenta o contato agonístico ou antagonístico entre os indivíduos (propiciando maior transmissibilidade dos parasitos). Já durante a estação seca a água existe disponível apenas em poços, lagoas e charcos que resistem. Isto acarreta uma convergência de animais para ingerir água nestes locais e uma consequente contaminação do ambiente peri-circundante à fonte d´água por fezes, urina e secreções dos animais a que nela chegam. Esta diversidade parasitária no local agrava a possibilidade de infecção cruzada entre espécimes e espécies. Enquanto o ambiente seco em geral diminui a contaminação ambiental, nestas regiões circunvizinhas à fonte d´água a maior umidade do solo permite uma sobrevivência das formas infectantes. Redes de transmissão parasitária inter-espécies podem se estabelecer nestas situações, para vantagem evolutiva do parasito, mas muitas vezes com prejuízo para o hospedeiro Já no segundo, aglomerações à volta da água remanescente, contaminação fecal e acúmulo mineral, são fatores comuns que alteram a ocorrência de doenças, infecciosas ou não (Amoroso et al. 2019).
O segundo aspecto é o da Qualidade da água. O quesito qualidade da água pode ser dividido em aspectos, referentes à sua qualidade biológicoa ou química.
4.1.1.4 Qualidade Biológica:
Que a água não tenha microorganismos em quantidade ou tipo suficiente para causar doença. Tais microorganismos podem ser advindos de contaminações do solo onde a água é estocada, de canos de condução da água quebrados que permitam a contaminação com matéria orgânica, de caixas d´água e cisternas sem tampa ou completament abertas, ou mesmo de açudes e poços artesianos que forneçam a água de bebida dos animais, situação comum em propriedades rurais. Qualquer Médico Veterinário que comece a trabalhar em um local novo deve pedir uma análise microbiológica da fonte da água ofertada aos animais para aferir o nível de contaminação com coliformes fecais totais e termotolerantes. Coletas e análises rotineiras (anuais pelo menos), são também recomendadas. Tais coletas e monitoramento agem como forma de vigilância sanitária na propriedade.
4.1.1.5 Qualidade Química:
A água completamente pura e desprovida de soluto de espécie alguma é uma artifício laboratorial. Em condições naturais mesmo a água mais cristalina e pura das montanhas tem minerais em solução. A questão reside na quantidade e qualidade dos minerais existentes nesta solução (López-Alonso 2012). Tais minerais são aproveitados pelos organismos animais e são importantes para a própria absorção da água. Entretanto, a ingestão através da água de bebida de quantidades exageradas de minerais comuns como Magnésio, Ferro ou Flúor podem comprometer a saúde dos animais, principalmente envolvendo o sistema renal, onde a formação de cálculos e desbalanço iônico são alguns dos problemas que podem surgir (Sengupta 2013). A concentração de minerais pode ser revelada por uma análise físico-química da água a ser ofertada aos animais, o que também é recomendado que seja feito rotineiramente (Gintam 2019).
Doenças advindas de contaminação através da água de bebida geralmente afetam muitos animais no plantel (não são problemas individuais), ou afetam instalações ou prédios específicos que compartilham a mesma fonte de água. Doenças trazidas pela água podem agravar-se gradativamente no plantel, por acumulação da toxina, agressor químico ou biológico. Ou surgirem abruptamente; neste último caso geralmente haverá algum evento anômalo acidental no local de criação que esteja ligado à origem da contaminação da água.
Animais criados em condições controladas (como laboratórios, canis, gatis, criações de pequenas aves, anfíbios e répteis) geralmente tem oferta de água comercial de alta qualidade, clorada, fluorada. Note que cloro e flúor podem estar em quantidades excessivas também para determinadas espécies aquáticas; neste caso menos é melhor. Tais fontes de água tem pequena probabilidade de contaminação biológica, mas caixas d´água sem tampa ou com animais mortos dentro, canos furados, contaminação cruzada com esgoto doméstico, todos são problemas que podem acontecer, mas devem ser antecipados pelo Médico Veterinário.
Entretanto, não é toda a criação animal que pode arcar com os custos altíssimos de utilizar água tratada. A água de bebida para animais de maior porte em geral vem de rios, poços artesianos, açudes, nascentes, todas fontes de água naturalmente sujeitas a uma maior probabilidade de contaminação. O Médico Veterinário deve estar pronto para sugerir ações que melhorem ao máximo a qualidade final da água, atuando desde a coleta de água, passando pelo transporte até a distribuição aos animais. Nestes casos, uma vigilância constante sobre a qualidade da água pagar-se-á em menor ocorrência de doenças e maior produtividade do plantel. No momento de planejar-se os sistemas de distribuição de água aos animais é interessante que haja como controlar-se o fluxo e quantidade de água ofertada, através de caixas d´água e válvulas de fluxo. Isto permitirá a viabilidade do estabelecimento de terapêuticas tendo como veículo para o fármaco a água de bebida. Esta estratégia facilita tremendamente a dosagem, a correta administração e distribuição entre os animais do fármaco, vacina, quimioterápico que o valha, sem perturbar os animais, com manejo humano mínimo e eficácia máxima. Novamente, a água com boa qualidade química e biológica é fundamental para o sucesso de tal estratégia terapêutica.
A água então pode comprometer a saúde dos animais tanto diretamente, por falta de ingestão ou por falta de qualidade (química), como indiretamente, por ser veículo para microorganismos e formas infectantes de parasitos em geral, que são ingeridos com a água e podem causar doença (Poppenga 2000). Assegurar água de bebida de qualidade para os animais é importante para o sucesso na mantença seja em condições de criação ou produção. Já para animais de vida livre o epidemiologista veterinário tem que ter em mente que a busca por acesso e ingestão da água vai moldar o comportamento do animal estudado e esta moldagem pode modificar a sua suscetibilidade a doenças.
4.1.2 Solo e Topografia
Desde a formação da crosta terrestre há 4 bilhões de anos atrás ela vem sendo modificada em sua constituição e forma. Tectonismo, quedas de meteoros, vulcanismo, erosão causada por água, poeira e vento todos eles modificaram a constituição do solo e da topografia terrestre. Estes processos de modificação geológica disponibilizam nutrientes minerais no solo, em diferentes quantidades e composições, de acordo com as rochas erodidas e os processos bio-geológicos locais. A estrutura do solo e sua composição química (assim como a disponibilidade de água, como visto acima) irão então favorecer apenas a microbiota e a vegetação que sejam capazes de sobreviver em tal solo. Assim, cada solo ira desenvolver uma determinada fitofisionomia, uma coleção vegetal (e de microorganismos), que se adapta e evolui em tais condições.
Cada tipo de solo se organiza de acordo com a topografia local (ou que venha a interferir localmente). Assim, nutrientes orgânicos e minerais, e parasitos, tendem a acumular-se na parte baixa das propriedades e/ou ambientes, devido ao efeito de carreamento da água. Este mesmo efeito de carreamento pode ser exercido pelo vento predominante, dependendo da topografia local. Nos dois casos devemos tentar alojar os animais com as idades mais jovens na regiões mais altas ou na posição da propriedade daonde vem o vento. Assim, o carreamento dos parasitos e/ou formas infectantes dar-se-á das idades mais jovens para as mais velhas, melhorando o controle de doenças.
Esta limitação de colonização biológica em cada tipo de solo tem por base o metabolismo de cada organismo. Se, e em que limites, ele pode utilizar os minerais de tal solo para executar suas funções metabólicas. É comum ouvirmos que determinada planta não cresce em solos pobres em Nitrogênio, ou ricos em Sódio, ou ao contrário, a presença de determinada planta indica excesso ou falta de determinados minerais. Uma determinada planta, por ser comumente imóvel, depende da coleção mineral contida no horizonte de solo limitado pela sua rizosfera (o espaço tridimensional envolvido pela raiz da planta), embora minerais carreados pela água possam ir e vir. Por conseguinte, cada consumidor primário (herbívoros em geral) só ingerirá como nutriente os minerais que ela tenha adquirido pela raiz, minerais estes limitados em disponibilidade pela coleção mineral local no tipo de solo. Esta composição diferenciada das plantas acarreta seleção de ingestão: diferentes animais ingerem preferencialmente plantas diferentes, em quantidades variáveis.
4.1.3 Atmosfera e Vento
Juntamente com a ação da água, a ação do vento com escultor de ambientes é importantíssima para lixiviação de rochas, pulverizando-as e permitindo que os microminerais possam ser carreados pelo próprio vento ou outros carreadores. A ação do vento é capaz de carrear tais microminerais por alguns poucos até milhares de quilômetros de distância, dependendo da intensidade do vento. A ciência reconhece hoje que milhares de toneladas de nutrientes minerais são depositados sobre a Amazônia e Europa, vindos da África, ou sobre a Índia, vindos da Ásia Setentrional, devido ao carreamento feito pelo vento em altas altitudes (Prospero, Glaccum, and Nees 1981). Este carreamento de nutrientes é importante fonte de nutrientes para as regiões que as recebem, mas o vento, além de nutrientes benéficos, pode carrear também plantas, animais, parasitos e suas formas infectantes, gases, fumos e vapores tóxicos, dispersando estes materiais a grandes distâncias (Hellberg and Chu 2016).
Plantas tóxicas podem ser levadas de fora para dentro de recintos de animais cativos, podendo causar doença neles. A ornamentação vegetal ao redor de recintos, estábulos, criadouros animais deve considerar que tais vegetais possam ser levados pelo vento para dentro de tais instalaçãoes, expondo os animais à ingestão; assim, plantas tóxicas devem ser evitadas em tais ornamentações. Animais terrestres e aquáticos podem ser sugados da superfície do solo pela força de tornados e furacões, sendo devolvidos à terra quando o vento enfraquece, às vezes à grandes distâncias. Existem vários relatos anedóticos de eventos em que “choveram” sobre o solo peixes, anfíbios, e até vacas e cavalos. Obviamente estes animais pode vir a ocasionar doenças nos locais que cairem, por apodrecimento das carcaças (que atraem também animais carniceiros, moscas e outros insetos), ou contaminação da água e comida por tais carcaças. Sem esquecer que toda a microbiota parasitária existente nos animais mortos potencialmente pode ser transmitida para a fauna da região de queda das carcaças. Tais eventos são incontroláveis, e o Médico Veterinário pode atuar apenas de forma paliativa.
Vetores invertebrados de doenças são particularmente suscetíveis à dispersão pelo vento, devido ao tamanho e peso pequenos. Assim, mosquitos que não costumam dispersar mais de 300-500 m através do vôo autônomo, podem ser carreados até 2 Km de distância pela ação do vento. Parasitos e suas formas infectantes (esporos, ovos ou formas vegetativas) podem invadir novos ambientes, aumentar sua velocidade de dispersão, infectar novos hospedeiros, todos efeitos de uma dispersão aumentada pelo vento (McManus 1988). Ambientes naturais fragmentados são particularmente prejudicados pelo aumento de entrada de parasitos causado pelo vento, já que há maior exposição à eles na borda do fragmento, tanto maior quanto maior for a proporção da área da borda em relação à area total do fragmento. Anteparos podem atenuar os efeitos do vento, devendo ser posicionados estrategicamente no local daonde vem o vento. Franjas de Eucaliptos, Pinheiros, ou outras espécies arbóreas, plantadas na propriedade podem ser utilizadas para criar tais anteparos. Note que um efeito indireto de controle de vento excessivo é a diminuição da desidratação da região sobre a qual o vento incide, melhorando a umidade da propriedade.
4.1.4 Clima
O último parâmetro abiótico acaba sendo a resultante de todas as variáveis acima, acrescidas de uma última: em qual latitude encontra-se o animal. Quanto mais ao pólos estiver posicionado o animal, menor será a incidência solar no local, ele tenderá a ser mais frio, e todos os seres vivos deste local devem adaptar-se a tal situação. Já para seres vivos localizados na linha do equador a incidência de sol é extrema, gerando um conjunto de demandas de saúde e bem estar completamente diferentes do animal localizado nas regiões polares. A esta questão básica, geolocalização e incidência solar, vão somar-se os outros fatores acima descritos (oferta de água, solo, topografia e vento) de forma a determinar o clima de uma região. Foge do escopo deste livro descrever a metodologia de determinação dos climas do mundo todo, mas o que nos interessa é que os animais vão ter sempre seu melhor desempenho fisiológico no clima a que foram adaptados evolutivamente, e que temos que a atender vários fatores e necessidades biológicas para que o animal fique em boas condições de bem estar, tendo boa saúde, e atinja sua produtividade máxima de crescimento, reprodução e longevidade.
No planejamento de instalações de criação animal deve sempre ser levado em suprema consideração o clima local. Sombreamento e resfriamento versus insolação e aquecimento, drenagem versus fornecimento de água, ventilação mínima versus ventilação assistida, todos são dilemas impostos por instalações localizadas em locais de climas diferentes. Erros crassos na criação animal já foram cometidos pela transferência pura e simples de um modelo de criação de locais frios para locais quentes e vice-versa. Assim, ter em mente no momento do planejamento das instalações qual espécie animal que vai ser criada, aptidão dela para o clima local, condições topográficas significantes, acesso a água, transporte, energia elétrica, e mesmo materiais de construção adequados à obra, todos são fatores fundamentais e que devem ser considerados para que tais animais possam ter desempenho fisiológico ótimo e que isto contribua para uma boa saúde.
O clima pode interferir na saúde dos animais devido às suas características intrínsecas, como temperatura, umidade, variações sazonais, extremos de temperatura e pluviosidade, ocorrência de ventos predominantes, intensos, ausência completa de vento, tempestades de chuva, neve e granizo, tornados, furacões, enfim, toda a gama de fatores que podem levar à comprometimentos da saúde dos animais. Doenças respiratórias são favorecidas por aglomeração dos animais, baixa renovação do ar ambiente, alta umidade, baixa temperatura, para citar alguns. Doenças originadas no aparelho digestório são favorecidas por alimento fermentado por chuvas intensas, modificações no pasto por enchentes, ou causadas por parasitos e suas formas infectantes que sejam carreados pelo vento intenso. Mortalidades de animais selvagens e de produção podem resultar de ventos intensos, tempestades elétricas, asfixia por fumaça levada pelo vento, nevascas e ondas de calor; fontes de radiação também podem ser dispersadas pelo vento. Em todos estes exemplos está a interferência ou alteração radical do ambiente do animal, ocasionado por algum evento climático. Fazer tal associação entre o evento e a doença é o desafio do Epizootiologista. Em algumas situações tal associação será fácil de ser feita (p.ex., 30 vacas mortas no pasto depois de uma tempestade de raios é fácil fazer a associação causal); em outras, a cuidadosa anotação de variáveis climáticas e das características da doença/morte dos animais (dia, hora, local, entre outras) vai permitir estabelecer uma correlação inicial, o que pode levar a eventualmente o estabelecimento de uma relação de causalidade entre tais eventos.
4.1.5 Considerações Finais
Vimos então que cada organismo biológico adapta-se e evolui de acordo com as condições de clima e geofisionomia local de um determinado ambiente, e que a saúde dos animais pode ser prejudicada por eventos e condições oriundos destes interferentes abióticos, que são incontroláveis para o animal. O ambiente é o primeiro vértice da tríade epidemiológica, sobre o qual se estabelecem as relações entre os outros seres vivos que nele consigam se estabelecer. Neste ambiente, microbiota, flora e fauna (termos genéricos) interagem na busca e conquista por território de onde retiram seus recursos nutricionais, conseguem abrigo e reproduzem. Desde a a obra fundamental de Charles Darwin está claro que é a competição, mais do que uma boa adaptação ambiental, o principal desafio que os seres vivos encontram no processo de sobrevivência: não basta ser bem adaptado, há que superar a concorrência. Os competidores mais adaptados reproduzem mais descendentes férteis. Este grau de adapatação (fenótipo) é expressão direta do genótipo de um animal, que contém a codificação gênica de tal bom competidor.
4.2 Fatores bióticos
Estes fatores são comumente responsivos aos fatores abióticos, tendo em vista que os organismos biológicos sofrem com as modificações das variáveis físicas. É claro que interferências de grande escala de organismos biológicos podem alterar determinadas variáveis físicas em escala global, como é o caso do corredor de umidade da Floresta Amazônica para a Mata Atlântica devido à evapotranspiração da floresta. Mas caso a cordilheira dos Andes subitamente ficasse plana, não haveria floresta amazÕnica que sustentasse aquele ecossistema, devido à radical modificação do fluxo hídrico que esta modificação causaria na região. Entretanto, Darwin foi o primeiro a caracterizar que a principal competição de qualquer ser vivo não era contra os elementos físicos, mas sim contra a concorrência biológico imposta por todos os outros organismos vivos também lutanto por recursos no mesmo ecossistema. Com esta ideia em mente, organizo os fatores bióticos naqueles ligados ou ao hospedeiro ou a toda a comunidade parasitária que nele se estabelece, considerando-se que determinados componentes desta comunidade parasitária podem ser mais prejudiciais ao hospedeiro do que outros.
4.2.1 Fatores ligados ao hospedeiro
Nosso principal foco de atenção neste livro são os vertebrados. Estão neste táxon nossos principais animais de interesse, seja na produção ou de companhia. Outros grupos de interesse podem ser eventualmente citados, todavia. Tendo isto em vista, considero fatores intrínsecos fundamentais, além de sua constituição genética, que interferem per se na competência imunológica de cada animal vertebrado, as seguintes variáveis:
- Sexo
- Idade
- Peso corporal
- Estado Nutricional
- Estado Reprodutivo
Estes aspectos são importantes a despeito da constituição genética ou da microbiota infectante de tal animal, porquê interferem diretamente na capacidade metabólica, competência imunológica e comportamento dos animais, modificando sua suscetibilidade às doenças. Note que em relação ao Estado Reprodutivo, como ele é uma resultante do sexo e da idade do animal, bem como de sua correta adaptação social. O mesmo acontece com Peso e Estado Nutricional, ambos estão ligados entre si e também são dependentes da adaptação social e ecológica do animal. Vamos então discutir a seguir os principais motivos porquê eles são relevantes para a análise da saúde dos vertebrados.
4.2.1.1 Constituição genética
Dois aspectos são cruciais na constituição genética de um determinado indivíduo que interferem na sua suscetibilidade às doenças: a variabilidade genética intrínseca de seu genoma e seu clado genética. Estes dois fatores podem interferir na produção metabólica que o indivíduo pode dedicar ao sistema imune, na diversidade e adequação dos clones de anticorpos e de seus epitopos aos desafios parasitários impostos, na capacidade de cicatrização dos tecidos, na tolerância aos efeitos dos parasitos e na resistência gerada pela imunidade inata a etapas do ciclo de vida dos parasitos.
O clado genético a que pertence determinado indivíduo também é um mapa que orienta o epidemiologista para a gama de potenciais suscetibilidades que tal animal pode ter frente ao histórico de parasitas e doenças que podem afetar o clado genético a que ele pertence. As grandes classes animais (peixes, anfíbios, répteis, aves e mamíferos) tem grau de compartilhamento diverso de parasitos, onde a temperatura corporal diferente é uma barreira sanitária natural. Parasitos de peixes, anfíbios e répteis são mais facilmente compartilháveis entre si, pelo metabolismo ectotérmico dos três grupos, do que com mamíferos e aves, já homeotérmicos e com temperatura corporal mais alta. Associado à esta questão metabólica ligada à classe animal, a própria linhagem evolutiva a que pertence o animal em questão também gera diferenças fenotípicas que podem reduzir a adaptabilidade de um determinado parasito a seu organismo, permitindo ou não o estabelecimento da infecção, e impedindo (ou não) a reprodução do parasito. A resistência de uma epiderme mais grossa à penetração de ectoparasitas é geneticamente determinada. Há resistência à infecção viral em determinado animal se ele não possuir epitopos de superfície que permitam a adsorção do vírion à membrana celular. A variabilidade do fenótipo molecular destes epitopos é dependente do clado genético do animal. Determinadas variantes genéticas podem tolerar melhor a infecção por hemoparasitas de vários taxa. Por essa linha de raciocínio, quanto mais geneticamente próximos duas espécies animais forem entre si maior é a possibilidade que parasitos possam infectar ambos, ou adaptar-se de uma para outra espécie. Estas barreiras moleculares ao salto de um parasito para um novo hospedeiro foram um dos mecanismos mais afetados pela globalização discutida no Capítulo 1, porquê aumentou subitamente o grau e a oportunidade de contato para um grande número de novos parasitos e novos hospedeiros. Mas barreiras genéticas ainda cumprem seu papel em evitar infecções por gerarem diferenças anatômicas, fisiológicas, imunes, únicas por vezes, mas que modificam a suscetibilidade do hospedeiro à doenças em geral, ou específicas.
No Capítulo 2 vimos que tolerância, resistência e resiliência são ramos distintos de fatores que afetam a atuação do hospedeiro como organimso biológico frente aos desafios impostos pelos parasitos (Best, White, and Boots 2008; Råberg, Graham, and Read 2009; Tadiri, Fussmann, and Scott 2021). Estes ramos têm gens distintos envolvidos na sua expressão final, e a seleção de tais genes é uma área importante da produção animal e da Medicina Veterinária e humana (Doeschl-Wilson and Kyriazakis 2012; Glass 2012; Kause and Ødegård 2012). Animais mais resistentes, com melhor tolerância à infecção, ou impérvios aos efeitos dela acarretam um efeito de redução do sucesso reprodutivo dos parasitos. Esta redução de sucesso reprodutivo do parasito torna as ações preventivas no nível secundário e terciário mais efetivas (vide Capítulo 11), e as ações de tratamento mais bem sucedidas.
Em termos de variabilidade genética, foi estabelecido que o mínimo desejável de variabilidade genética de uma população animal seja cerca de 95%, ou seja, um parentesco (inbreeding) entre os indivíduos da população menor do que 5% (Frankham 1994; Groeneveld et al. 2010). Este é, por exemplo, um dos critérios para inclusão de populações ou espécies de animais silvestres de vida livre na categoria de criticamente ameaçada de extinção: tal tamanho de população animal não apresenta, se mantida nas condições atuais, viabilidade para manter 95% de variabilidade genética nos próximos cem anos, devido ao conjunto de pressões ambientais que cercam-na (Padrões e Petições da UICN 2022). É claro que em situações de cativeiro de animais silvestres, com animais de produção, ou na criação de animais de laboratório é desejável que mantenhamos tal nível de variabilidade genética, a não ser quando a homozigosidade seja um modelo experimental válido, como nas linhagens de camundongos consanguíneos. Em todos estes casos é mister um controle de reprodução e de pedigree dos animais, para que seja assegurado o nível de variabilidade genética desejado.
A falta de variabilidade genética, homozigosidade portanto, leva a efeitos deletérios de cunho imunológico, fisiológicos, anatômico e cognitivos à saúde dos animais (Lacy 1997; Reed and Frankham 2003). Todo Médico Veterinário deve tentar manter a variabilidade genética dos animais sob sua responsabilidade dentro de limites razoáveis, através da introdução de novos animais (ou sêmen, óvulos congelados), rotação de reprodutores, manutenção de tamanho de população adequado, entre outras ações (W. G. Hill 2000). Populações animais domésticas e de produção são particularmente propensas a homozigosidade (Makino et al. 2018), e devem ser propriamente manejadas.
4.2.1.2 Sexo
Um aspecto que pode ser observado na Seleção Natural é que foram favorecidos os métodos de reprodução que tendem a gerar variabilidade genética da progênie de forma controlada, e isso é atendido de diversas formas. A reprodução sexuada é uma destas estratégias de sucesso evolutivo entre os animais pluricelulares; raras são os clados em que isso não acontece. Desde a cópia do DNA nas células de linhagem, passando pela meiose e crossing-over, e culminando na reprodução sexuada em suas diversas variantes, todas estas são instâncias em que, de forma mais ou menos controlada, variabilidade genética tende a ser produzida (Ellegren and Galtier 2016). Além disso o acaso interfere, através de mutações, deleções e inserções de genes, ou imigrações de genes animais (Maia et al. 2021). A estratégia reprodutiva sexuada separa geneticamente a espécie em dois sexos, aqui denominados macho e fêmea, os quais possuem características distintas em aspectos de morfologia e fisiologia que os distinguem geralmente em tamanho corporal, colorido, comportamento, em suma várias características animais são diferentes nos machos e fêmeas (Adkins-Regan 2007; Hau 2007). Estes genomas com os mesmos genes, mas alelos potencialmente diferentes são misturados quasi-aleatoriamente no ato reprodutivo, gerando uma progênie que pode ser de qualquer um dos sexos (Maia et al. 2021).
Dentre os vertebrados especificamente, um aspecto ligado às diferenças de qualidade e intensidade da resposta imune difere muito entre machos e fêmeas: a maior secreção de testosterona pelos machos a partir de certa idade, no que seria o período da puberdade da espécie. Sabe-se que os caracteres reprodutivos secundários dos machos (colorido exuberante, cornos, chifres, esporões, cristas, barbelas, maior tamanho corporal, entre outros) são exibidos em resposta à testosterona secretada pelo animal, e que tal secreção depende de raça/linhagem, qualidade da alimentação, estado sanitário, idade e qualidade do ambiente de alojamento do animal, principalmente (Hau 2007). A partir da caracterização da existência de um mecanismo fisiológico de retro-alimentação positiva entre a secreção de testosterona e a de cortisol, foi sendo confirmado em várias espécies de diferentes classes de vertebrados, um período de queda de imunidade inata e adquirida por parte dos machos, justamente durante tal puberdade. Tal queda de imunidade nos machos também pode ser vista em idade mais avançada, geralmente nos períodos de formação de harém e/ou disputa por fêmeas por outros machos (Kempenaers, Peters, and Foerster 2007). A explicação deste fenômeno pode ser metabólica, já que qualquer processo fisiológico que requer maior crescimento de tamanho corporal, qualidade de empenamento ou pelagem, crescimento de apêndices reprodutivos (como cornos, chifres, cristas, barbelas), requer obrigatoriamente maior gasto de energia, gasto este que, no orçamento metabólico que qualquer animal tem que manter entre ingestão e dispêndio diário de energia e nutrientes, tem que ser coberto pelo menor investimento em energia no sistema imune, que é naturalmente caro em termos de requerimento de energia e nutrientes (S. L. Klein 2004).
As fêmeas também passam por períodos de queda de imunidade fisiológica durante a época e/ou idade reprodutivas, fases de alta demanda fisiológica e de produção de energia para a fêmea. Sabe-se há muito tempo, primeiro por experiências na criação animal, e mais recentemente por avaliação imune estrita, que as fêmeas tem uma queda de imunidade peri-puerperal. Nos mamíferos tal queda ocorre no terço final do período gestacional e no terço inicial do período de aleitamento, fases em que a fêmea tem que prover, às custas do seu próprio metabolismo, tanto o crescimento em massa corporal do feto em seu desenvolvimento uterino final, como a carga calórica que o leite tem que fornecer ao neonato para seu início de desenvolvimento, até que ele atinja a ingestão de alimento por si só. Nas aves fêmeas tal fase de queda de imunidade ocorre no final da preparação dos ovos para a postura e no período inicial de cuidados com o nidícola (peixes, anfíbios e répteis provavelmente tem fases semelhantes), também fases que demandam alto investimento em produção metabólica pela fêmea (Clutton-Brock 2009; Sabra L. Klein and Flanagan 2016).
Em termos evolutivos, a retro-alimentação positiva entre testosterona e cortisol, com consequente expressão dos caracteres reprodutivos secundários nos machos, resulta que tais caracteres são indicadores honestos de qualidade genética para a fêmea. Se aquele macho tem um pescoço mais grosso que os outros, é maior, mais vistoso e colorido, ou é superior em qualquer aspecto morfológico ou comportamental que seja, apesar de proporcionalmente imunossuprimido (ele não ficou doente, em suma), isto é um indicador honesto para a fêmea que ele tem qualidade genética. Tal qualidade genética do macho reverteria-se diretamente na melhora da fertilidade do casal, por indicar que ele é capaz de defender um território de alimentação adequado, protegê-la durante o perído reprodutivo e alimentar a progênie, além de transmitir para esta última um genoma bem adaptado aos desafios do meio (Negro et al. 2010).
Outro aspecto deve ser chamado à atenção em termos de constituição do sistema imune de machos e fêmeas: as células de linhagem espermática dos machos são mantidas isoladas do contato com o sistema imune do próprio organismo, enquanto as células de linhagem da fêmeas tem contínuo contato com o millieu imune, sofrendo os mesmos estímulos agonísticos causados pelas citocinas, anticorpos e outros peptídeos trazidos pelo sangue como qualquer outra célula orgânica. Desta forma, podemos considerar que as células de linhagem espermática dos machos estão como que congeladas no tempo molecular da época em que seu ovócito secundário foi fecundado. Já tal ovócito secundário fecundado carrega consigo toda a cena molecular a que a fêmea está submetida, os mesmos estímulos moleculares a que aludimos o espermatozóide nunca será submetido (Kierszenbaum and Tres 2019). Tal arranjo evolutivo permite que, a cada geração ocorra uma mistura entre o material genético do macho, que é um registro genético “congelado” do que foi bem sucedido evolutivamente na geração anterior, com o material genético E célula da fêmea, ambos sob o efeito da cena molecular atual, que representa todos os fenômenos moleculares a que ela foi submetida em sua vida até aquele momento (Pennell, Galligan, and Fish 2012). Este tipo de diferença ontogênica na constituição e produção das células de linhagem reprodutiva altera a resposta de machos e fêmeas frente aos desafios parasitários de formas ainda não completamente compreendidas (Fink and Klein 2018).
4.2.1.3 Idade
Todo animal ao nascer tem como mecanismos de defesa básicos sua imunidade inata e o pool de anticorpos conferidos pela mãe. No caso da imunidade inata, macrófagos, células natural killer, células dendríticas, todos são células efetoras que executam principalmente fagocitose e apresentação de antígenos para as células responsáveis pela ramo humoral da defesa imune (Iwasaki and Medzhitov 2010). Já no caso dos anticorpos maternais, estes são conferidos segundo o pool circulante possuído pela mãe, permitindo ao recém-nascido ativar seus primeiros Linfócitos B circulantes; os anticorpos maternais defendem o recém nascido durante o período em que os anticorpos próprios aumentam gradativamente. Os anticorpos maternais desaparecem durante os primeiros 30-40 dias de vida na maior parte das espécies de vertebrados. Tomadas as diferenças entre animais jovens e adultos em conjunto, animais jovens tem menos células de defesa, estas são menos responsivas, as respostas são mais lentas e menos específicas; o resultado é que frente á maior parte dos desafios parasitários, animais jovens são mais suscetíveis do que adultos (Marchant and Kollmann 2015).
Já na outra extremidade da linha de vida de qualquer ser vivo, vertebrados que atinjam a senilidade também tem sua imunidade comprometida, por outros motivos. Em animais mais velhos a capacidade de produção de energia diminui, e também a energia capaz de ser direcionado para o sistema imune, tornando-o menos capaz de responder a desafios parasitários intensos. Novamente, genomas bem adaptados às condições de desafio do ambiente podem manter bons níveis de saúde em idades avançadas, onde animais da mesma espécie já demonstram sinais de debilidade imunológica e orgânica. Outro aspecto importante na maior suscetibilidade à doenças em animais mais velhos é a própria gradual inadequação do sistema imunológico à evolução constante da microbiota no ambiente onde ele vive. A cada 24 horas uma nova geração de bactérias se forma, uma partícula viral produz milhares de cópias de si mesma em uma única célula infectada (cada uma com potencial mutações), prototozoários multiplicam-se exponencialmente a cada geração no hospedeiro, enfim há uma mudança de constituição genética gradual da microbiota que infecta um animal que se realiza durante o tempo de vida dele. Entretanto, o hospedeiro encontra-se congelado geneticamente no momento de sua concepção, que pode variar de poucos meses no passado até a várias décadas, ou até mesmo centenas de anos, como no caso de algumas espécies tremendamente longevas (répteis, p.ex.). Em determinado momento há um descompasso entre o tipo de resposta humoral que tal hospedeiro pode apresentar e a qualidade do desafio parasitário a ele imposto. O animal começa a tornar-se suscetível a microorganismos até mesmo de sua microbiota normal e sucumbe (Dowling and Levy 2014).
Em resumo são mais suscetíveis a doenças animais em idades muito jovens, principalmente durante o período inicial de produção de linfócitos T e B e maturação de seu sistema imune, e animais muito velhos, devido à debilidade imune e orgânica gradativa que eles desenvolvem (Grubeck-Loebenstein 1997; Weiskopf, Weinberger, and Grubeck-Loebenstein 2009). Notem que toda regra em biologia pode ser seguida de vários exemplos em contrário e isto não é falso para a suscetibilidade devido à idade (Thaiss et al. 2016; Martino et al. 2022). Determinados agentes parasitários não são capazes de infectar animais muito jovens ou não causam malefício a animais muito velhos, não necessariamente por eles estarem fora do grupo de risco (como no caso de doenças venéreas), mas por não apresentarem as condições fisiológicas e mesmo epitopos de ligação apropriados para tais agentes parasitários (Wassom 1993; Sadighi Akha 2018).
4.2.1.4 Peso corporal
Ter um tamanho corporal maior é vantajoso para a maior parte dos seres vivos, e crescer em tamanho é a tendência de espécies animais em condições de boa adaptação ambiental. Mas para crescer é necessário tempo, ou seja uma longevidade maior também é vista em animais maiores, em média. Isto pode ser bem visto na produção animal onde animais maiores são obtidos gradativamente à medida que melhoram as condições de bem-estar e qualidade na criação animal; galinhas, suínos, bovinos são abatidos hoje em dia com rendimentos de carcaça bem maiores do que há cem anos atrás. Estes dois fatores, longevidade e tamanho corporal maiores alteram a suscetibilidade à doenças destes indivíduos ou espécies por motivos diferentes.
Primeiro, quanto mais longevo um animal maior seu tempo de exposição aos parasitos e mais tempo estes tem para causar alterações fisiológicas dignas de nota em tal hospedeiro, principalmente em parasitos com ciclo de vida mais longo. Segundo, quanto maior o hospedeiro em tamanho corporal, maior é o “ônibus parasitário” que ele representa, ou seja maior é a diversidade ou a carga parasitária que ele pode albergar. Isto também reflete as diferenças de quantidade de metabolização de energia diária que existem entre um animal pequeno e um de grande tamanho, onde o último adquire e metaboliza uma quantidade maior de energia, sendo capaz também de fornecer oportunidades de ganho energético a um número e diversidade maior de parasitas. A própria maior ingestão de alimento para fornecer tal energia também aumenta a possibilidade de ingestão de maior diversidade e quantidade de parasitos e suas formas infectantes, além do animal se movimentar mais por ter que percorrer um território maior que o alimente.
4.2.1.5 Estado Nutricional
“Saúde entra pela boca.” Anônimo
O reconhecimento de que o alimento faz diferença para uma boa saúde é reconhecida há muito tempo, embora só no século passado tenham sido caracterizados os diferentes componentes nutricionais que devem constituir um “prato de comida” adequado para qualquer animal. Para definir tal prato adequado, um Médico Veterinário deve conhecer os seguintes aspectos do animal:
- Sua classe animal e espécie: Com estas duas informações podemos determinar em qual classe de taxa metabólica este animal estará, e qual sua história evolutiva de consumo de alimento. Digamos, se o animal é carnívoro, frugívoro, de pastejo, piscívoro, onívoro, ou outra entre as diversas categorias possíveis.
- Seu peso corporal: Necessário para calcularmos sua taxa metabólica basal e assim derivar outros quantis de energia necessários. Acurácia na medida é necessária, variações de precisões de não mais de 10% são recomendáveis na aferição do peso dos animais (se o animal tem 1 Kg real eu posso estar medindo algo entre 900g e 1100g).
- Sua fase de vida e estado sanitário: O consumo de energia calculado pelo peso do animal deve ser ajustado para outras demandas de energia que são dependentes da idade e/ou fase de vida do animal, como crescimento e reprodução, e a demanda gerada pelo sistema imune. Animais adultos não reprodutivos podem dedicar mais energia ao sistema imune, melhorando a resposta como um todo. Animais em crescimento ou reprodução, tem limites mais rígidos para dedicação de tal quantidade de energia, acarretando imunossupressão. A boa alimentação nestas fases de maior exigência calórica e nutricional permite uma atenuação destes efeitos, enquanto deficiências alimentares e nutricionais acarretam agravamento dos quadros infecciosos.
Definidas as demandas de um determinado animal, estas devem ser atendidas pela alimentação. Mas por alimentação adequada, três aspectos devem ser atendidos por tais demandas:
Quantidade de energia adequada: Definida com base no cálculo da taxa metabólica basal, levando em consideração tamanho corporal e classe do animal em questão, ajustada para as demandas adicionais, como manutenção, sistema imune, reprodução e crescimento.
Substrato de energia de origem adequada: Definido com base na história evolutiva alimentar da espécie do animal em questão. Se o animal é carnívoro, ele deve ter sua principal fonte de de energia oriunda de proteína animal. Se o animal é insetívoro, ele deve ter sua principal fonte de de energia oriunda de invertebrados. Se o animal é herbívoro de pastejo, ele deve ingerir sua principal fonte de energia de gramíneas de boa qualidade. Se o animal é frugívoro, sua principal fonte de energia devem ser carbohidratos (hexoses em geral). E assim sucessivamente, sob o arrazoado de que todo o sistema digestório do animal está otimamente ajustado para absorção e metabolização da fonte de energia a que ele foi evolutivamente adaptado.
Microminerais em quantidade e diversidade adequadas: na tabela periódica da Figura 4.4 foram assinalados os átomos que já foram caracterizados como participantes de algum sistema enzimático ou metabólico biológico. Pode-se notar que alguns são bem conhecidos como íons importantes para a dinâmica fisio-metabólica de um vertebrado, outros que eram conhecidos como elementos-traço, hoje são reconhecidos participantes em processos orgânicos.
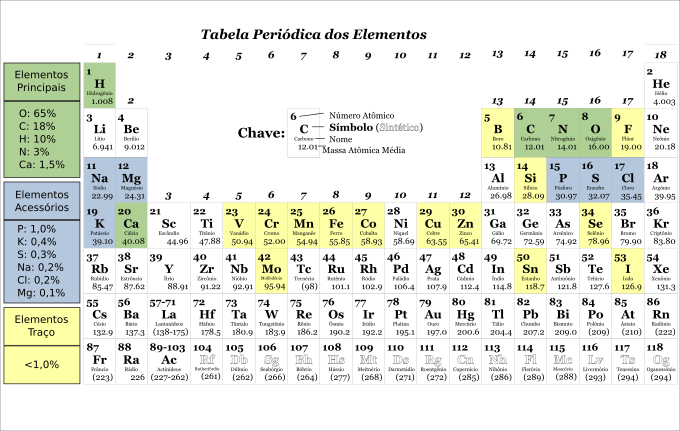
Figura 4.4: Tabela periódica ressaltando os elementos químicos que constituem usualmente os vertebrados, bem como a proporção de contribuição de cada um.
Cada organismo biológico terá então uma necessidade de absorção de energia (de fontes específicas) e microminerais dependendo de suas características fisiológicas, evolutivamente adquiridas. Animais marinhos, por exemplo, muitas vezes possuem glândulas específicas para a excreção de sal marinho (NaCl), enquanto para animais terrestres a aquisição de sal é muitas vezes difícil. Tais diferenças fisiológicas geram tolerâncias diferentes aos níveis mínimos e máximos de ingestão de microminerais. O quê é muito para uns pode ser pouco para outros. Tais níveis mínimos e máximos de ingestão de microminerais foram gradativamente sendo estabelecidos, durante o século XX principalmente, para os animais de produção primeiro (bovinos, equinos, galinhas; Suttle (2010)) e posteriormente para animais domésticos (cães, gatos; Council et al. (2006)). Hoje nossa compreensão evoluiu para que administremos níveis maiores ou menores de determinados minerais dependendo da condição de criação do animal, idade, sexo, por reconhecermos que tais fatores geram demandas diferentes por determinados microminerais. O problema é saber tais níveis mínimos e máximos de ingestão para a miríade de espécies animais que compõem um ambiente natural. A comparação entre espécies estabulados e de vida livre é uma alternativa para inferir níveis adequados destes minerais (Babicz and Kasprzyk 2019; Sach et al. 2020).
Em diversas situações, deficiências de microminerais estão associadas a efeitos deletérios à saúde, tanto de forma direta como indireta, dependendo do mineral envolvido. Seja Sódio, Cálcio, ou Ferro, para citar aqueles de maior “renome”, ou seja Cobalto, Enxofre e Selênio, não tão citados (entre outros), todos são microminerais que participam de processos enzimáticos para a produção de energia, anticorpos e controle de radicais oxidativos, todos são relevantes e devem ser ingeridos em forma e quantidades adequadas. Embora eu esteja ressaltando problemas de origem carencial de minerais, o excesso de ingestão de minerais como Sódio, Flúor, Cálcio, entre outros, também pode levar a morbidades graves. A produção animal melhorou muito em produtividade quando o “premix mineral” passou a ser misturado às rações, de forma a corrigir e suprir as eventuais deficiências minerais que os animais pudessem vir a ter devido ao tipo de ingesta que ingerissem.
Em última instância, a mobilidade animal torna-se uma estratégia para aumentar a diversidade de ingesta que eles possam adquirir, vindo de plantas de diferentes solos, diminuindo a probabilidade de que haja carência de minerais na dieta. Este problema de aquisição de nutrientes é limitado pela escala de tamanho do animal. Um roedor de 35 g tem um território de forrageamento proporcional a seu tamanho diminuto; assim, sua ingesta de minerais é limitada por quais plantas crescem no solo limitado por seu território. Quanto maior o animal menor este problema se torna, porquê o território de forrageamento cresce proporcionalmente ao tamanho corporal, diminuindo a probabilidade de carência de minerais por aumento de oferta. Animais consumidores secundários (carnívoros) sofrem muito pouco com carências minerais em natureza, por ingerirem outros animais, que por sua vez já “coletaram” nutrientes e minerais para seu próprio organismo. Um efeito de bioacumulação de minerais ocorre, concentrando-os finalmente nos consumidores terciários.
Em resumo, devemos considerar estes aspectos:
- variabilidade de disponbilidade de minerais no solo e/ou nas plantas em cada ambiente pode gerar problemas carenciais ou de excesso de ingestão nos animais que nele habitam;
- A capacidade de adquirir microminerais no ambiente é diretamente proporcional ao tamanho do animal; quanto maior, maior o território, menor a probabilidade de excessos ou carências;
- A demanda por microminerais é fundamental para uma boa saúde e pode variar tremendamente em cada organismo animal.
Assegure-se que o problema de saúde que você esteja estudando não é originado ou agravado por dietas ricas ou pobres em determinados microminerais. Análises da composição química do solo (e da água que passa por tal solo), análise de plantas incluídas na dieta, extrapolações a partir de dados de outros animais, todos são estratégias para abordar tal pergunta.
4.2.2 Fatores ligados à comunidade parasitária
Eu discorri durante este capítulo sobre os fatores relacionados ao ambiente e ao hospedeiro que modificam a suscetibilidade deste às doenças. Agora é chegada o momento de discutir o parasito, ou agente etiológico putativo, ou patógeno, ou seja, um determinado microorganismo que, segundo seus efeitos sobre o hospedeiro (ou comunidade de espécies hospedeira), pode ou está causando a doença. Como vimos no Prefácio, a conceituação estabelecida no início da Teoria dos Germes era de uma abordagem epizootiológica “Parasita causa Doença”. Esta inequívoca afirmação foi sendo lapidade e melhorada com a melhor compreensão do início deste século para uma abordagem “Parasitos podem causar Doença, dependendo de muitos fatores” (Poulin 2004). Destes muitos fatores, a composição da própria comunidade parasitária que infecta o hospedeiro, em termos de diversidade (de espécies, levadas em consideração no nível genômico) e abundância (de cada destas espécies, tanto absoluta como relativamente à população total) faz diferença no resultado final de comprometimento ou não da saúde do hospedeiro (Ezenwa et al. 2010; Johnson and Hoverman 2012; Johnson et al. 2013). De fato, vários parasitos foram identificados como benéficos para seus hospedeiros, o qual sem aqueles não consegue manter bons níveis de saúde (McKay 2006; Barry 2022). Evoluções no estudo da imunologia deixam claro que mesmo a ordem de infecção a que o animal recém-nascido é submetido modifica seu perfil imunológico (Cox 2001; Karvonen, Jokela, and Laine 2019). O próprio sucesso ambiental do hospedeiro, aumentando seus números, acarreta modificações na estrutura da comunidade parasitária (Arneberg et al. 1998).
Entretanto, esta avenida de incógnitas imunológicas, ecológicas, parasitológicas, genéticas, entre outros ramos da ciência, ainda precisa de maior corpo experimental sob esta nova mentalidade para evoluirmos na compreensão da complexa relação parasito-hospedeiro. Este maior número de informações necessários não será construído se continuarmos a estudar doença em animais sem levarmos em conta o maior número de informações possíveis sobre a comunidade parasitária que o infecta, prestando atenção mais central a parasitos sabidamente capazes de causar doença clínica, mas sem esquecer que parasitos coadjuvantes certamente participam para definir a direção da resultante final: doença ou saúde. Assim, a mensagem final para quando o Médico Veterinário abordar uma investigação epizootiológica é: “Atenção com as co-infecções!”.
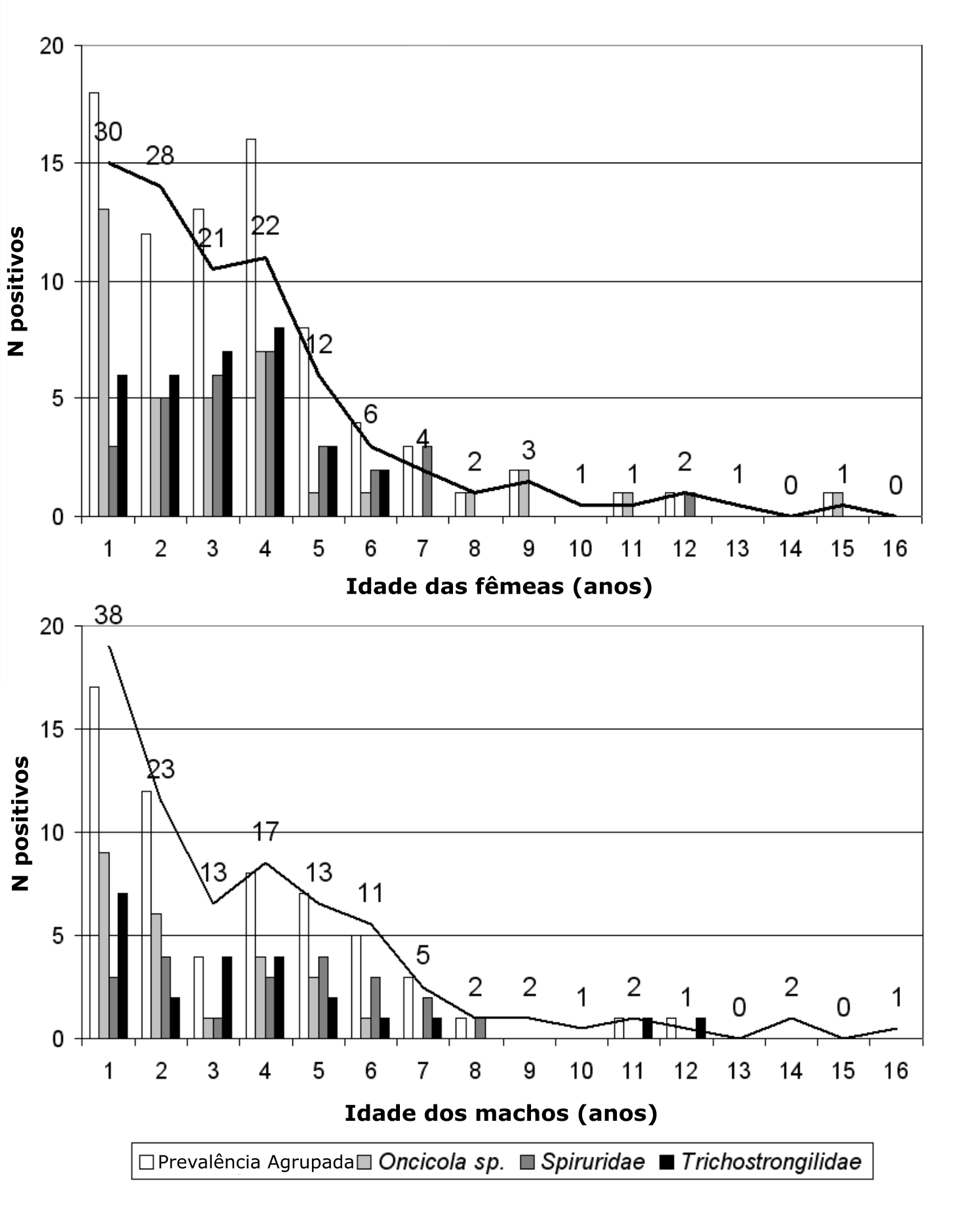
Figura 4.5: Variação com a idade da prevalência de 3 taxa de helmintos entéricos de Micos leões dourados, divididos pelo sexo do indivíduo. Note-se as variações em frequência da infecção entre os sexos, em dependência com fatores ontogênicos (puberdade), reprodutivos (gestação ou macho reprodutor) e com a própria espécie de helminto infectante. Uma análise das fontes de variação da prevalência pode ser vista no artigo original. Traduzido e adaptado de (Rafael V. Monteiro, Dietz, Beck, et al. 2007).