Capítulo 6 Investigação Epizootiológica
Como já visto no Capítulo 2, a Epidemiologia Clássica considerava com mais ênfase as características ligadas ao parasito, porquê desde o advento da Teoria dos Germes havia a ideia de que o parasito é o culpado. Através do século passado e início deste houve uma evolução nesta mentalidade, e a doença agora é vista como uma resultante de fatores ligados não só ao hospedeiro (já discutidos no Capítulo 4) ou ao ambiente mas também à comunidade parasitária, cujos efeitos benéficos e deletérios sobre o hospedeiro e seu sistema imune modificam sua suscetibilidade às doenças. Portanto, quando da execução de uma investigação epidemiológica, precisamos fazer cinco perguntas, cujas respostas nos permitem saber aspectos importantes relativos à doença em questão. Estas perguntas orientam a investigação epidemiológica e podem ser respondidas simultaneamente ou gradativamente:
- Quem são os hospedeiros afetados pela doença?
- Quando a doença ocorre?
- Onde a doença ocorre?
- Como e Porquê a doença ocorre?
- Quem é(são) o(s) agente(s) etiológico(s) envolvido(s)?
6.1 Quem são os hospedeiros afetados pela doença?
Normalmente uma investigação epidemiológica começa porquê alguma espécie(s) animal(is) ou população(ões) está apresentando alguma visível modificação de comportamento, física ou clínica que está acarretando a doença, morte ou perda de produtividade atlética e/ou reprodutiva nestes animais de interesse. O esforço de detecção, o interesse público e o senso de urgência na resolução do problema é aumentado quando os efeitos na população animal são agudos e graves, em oposição às doenças crônicas e brandas, que demoram para serem detectadas e diagnosticadas, por passarem desapercebidas. O local em que os animais estão sendo afetados também interfere na detecção da doença, já que animais em ecossistemas ou regiões pouco sujeitas a algum tipo de vigilância epizootiológica ou com baixa presença humana, tendem escapar à atenção tanto do público leigo como do técnico especializado, retardando a detecção da alteração mórbida no curso do tempo. A própria relação do homem com a espécie afetada também modifica a detecção da doença: doenças que afetem animais domésticos ou de produção geram um interesse de pesquisa e demanda pública de pesquisa muito maior do que doenças que afetem camundongos, ratos, urubus ou outros animais silvestres ou sinantrópicos que são considerados nocivos à espécie humana. Entretanto, tais doenças eventualmente podem espalhar-se para os animais mais “nobres”; a necessidade de pesquisa e investigação é relevante em qualquer situação.
O epizootiologista deve ter o cuidado de anotar todo e qualque animal que demonstre os sinais clínicos de interesse. Esta anotação deve fornecer o maior número possível de informações sobre as características intrínsecas do indivíduo acometido, em particular os fatores de interesse biológico que ressaltamos no capítulo 4: Espécie/raça/linhagem, Sexo, Idade (ou faixa etária), Peso Corporal e Estado Reprodutivo. É certo que para determinadas condições ou animais, tal colheita de dados é fácil e rápida, como no caso de animais de cativeiro, laboratório ou produção confinada. Muitas vezes os dados acima citados já são controlados naturalmente durante o processo de criação animal, facilitando a colheita de dados.
Tal não é verdade no caso de animais de vida livre, selvagens ou não, ou animais de produção criados de forma semi- ou extensiva. Nestes casos, a informação mais fácil de ser conseguida é a da espécie afetada, enquanto que para Sexo, Peso Corporal, Estado Reprodutivo e a colheita de material biológico para análises clínicas, é necessário a captura, e muitas vezes anestesia do animal, o que complica e encarece o procedimento de colheita de informações. Mesmo capturado, para a informação de Idade muitas vezes só é possível dividir-se os animais em jovens e adultos, o que reduz tremendamente a qualidade de tal informação. Entretanto, todo esforço deve ser empregado para que tais informações sejam conseguidas, em ordem de orientar adequadamente a investigação. Quanto melhores os dados, melhor o resultado final e mais fidedignas serão as conclusões sobre o processo mórbido em análise.
Cuidado deve ser tomado nesta etapa para que não haja erros de:
Identificação de espécie animal: determinadas espécies animais podem ser facilmente confundidas entre si (pelo técnico sem treinamento). Espécies denominadas crípticas muitas vezes só são diferenciadas da outra por exames moleculares, que mostram distâncias genéticas às vezes consideráveis para animais muito similares entre si. A mistura de dados destas espécies pode gerar um resultado final completamente inadequado para ambas as espécies confundidas. Atualmente as ferramentas moleculares permitem a caracterização inequívoca de espécies diferentes, e tal ferramentas devem ser usadas quando existir a possiblidade de confusão entre espécies. O testemunho da espécie em nossos dias deve ser preferencialmente molecular, e não meramente o depósito de carcaças representativas de alguns animais capturados, o que era o padrão (usado com sucesso variável) durante os anos passados.
Determinação de sexo: embora algumas espécies de vertebrados apresentem claro dimorfismo sexual, para outras espécies a aparência externa de machos e fêmeas é indistinguível ao olhar humano, com ou sem treinamento. Exames complementares tem que ser realizados sobre material biológico obtido do indivíduo para confirmar seu sexo. Estes exames podem envolver citogenética (caracterização dos cromossomos sexuais), colorações citológicas especiais (permitem evidenciar o corpúsculo de Barr - cromatina sexual), endoscopia (visualização direta dos órgaõs reprodutivos dos animais), sorológicas (permitem também o uso de excreções e secreções orgânicas) e, principalmente, técnicas moleculares, que devem sempre ser adequadamente validados previamente ao uso corriqueiro da técnica.
Aferição de Peso: Métodos diretos (balanças em geral) e indiretos (geralmente com o uso de correlações entre medidas corporais lineares e o peso corporal) podem ser usadas corriqueiramente para aferiação de peso corporal dos animais estudados. Em casos de animais muito pequenos cuidado deve ser tomado para que a precisão da balança seja adequada, em torno de 5% do peso do animal. Ou seja, se o animal pesar 100 gramas, a balança deve ser capaz de registrar variações de 5 em 5 gramas; se o animal pesar 20 Kg, a balança deve poder registrar intervalos mínimos de 1 Kg, e assim sucessivamente.
Estado reprodutivo: Nos casos de animais com reprodução sazonalmente demarcada, um pesquisador pode saber a época do ano em que os animais não estarão reproduzindo. Mas dentro da época reprodutiva, principalmente nas fêmeas, pode ser difícil que ele tenha certeza de uma prenhez. A inspeção visual pode auxiliar na determinação de prenhez, mas geralmente apenas no fim do período gestacional, onde os sinais evidentes e o comportamento diferente da fêmea ajudam. Técnicas não-invasivas podem ser usadas, como a análise de hormônios fecais. Mas muitas das técnicas viáveis de deterimnação de estágio de prenhez são invasivas, como palpação, colpocitologia, radiografia, ultrassom e endoscopia. Todas estas exigem a captura do animal, com os fatores complicadores que isto acarreta dependendo da espécie ou população trabalhada.
6.1.1 Diferentes papéis do hospedeiro na dispersão das doenças
Tendo em vista que todos os organismos biológicos existentes no planeta são diferentes geneticamente entre si (Muraille 2018), é nada mais do que natural que o fenômeno parasitismo envolva um espectro de adaptações entre um dado indivíduo parasito e o organismo hospedeiro infectado. Este espectro de adaptações é gerado não só pelas diferenças genéticas (e portanto de fenótipo) entre parasito e hospedeiro, mas também pelas interações entre si de cada um dos parasitos componentes da comunidade parasitária que o hospedeiro alberga, que vão sendo estabelecidas no curso do processo evolutivo (Papkou et al. 2016). Portanto, a relação parasito-hospedeiro individualmente estabelecida é modulada substancialmente pelas cooperações e competições entre parasitos individuais e suas populações (Rigaud, Perrot-Minnot, and Brown 2010). O primeiro antibiótico desenvolvido pela ciência, a penicilina, foi justamente uma proteína produzida por um fungo para manter à distância a competição bacteriana (Gaynes 2017). Biofilmes bacterianos estabelecem-se e cooperam entre si em orgãos intestinais e resistem à competição de outros parasitos localmente (Hall-Stoodley, Costerton, and Stoodley 2004). Tais mecanismos foram estabelecidos pela Seleção Natural no curso da evolução de parasitos e hospedeiros e, em última instância, ofereceram vantagem competitiva aos parasitos e hospedeiros.
O hospedeiro deve então ser compreendido como um ecossistema em si, com seu ambiente característico, que varia dependendo do compartimento orgânico onde o parasito se encontra, ambiente que determina as regras de engajamento entre as populações de parasitos lá estabelecidos e o hospedeiro (hosp como ecossistema). Neste ecossistema o número de variáveis que influencia a relação parasitos-hospedeiro é muito grande, e em grande parte desconhecido ou não avaliado pelos métodos científicos atuais (Mardahl, Borup, and Nejsum 2019; Forestiero 2022). Tanto é o caso que a caracterização de padrões globais na relação parasita-hospedeiro ainda é restrita a poucas variáveis (Rigaud, Perrot-Minnot, and Brown 2010).
Para tentar categorizar estas diferenças de sucesso de infecção de parasitos em diferentes hospedeiros foram primeiro caracterizadas espécies animais que eram “reservatórios naturais” de determinados parasitos, e a presença de tais espécies seria suficiente para manter uma população parasitária viável em um determinado ambiente. A presença de outros animais era algo irrelevante, tendo em vista os animais que eram “reservatório natural”. Mas com a evolução da parasitologia e epidemiologia desde o fim do século passado, este modelo dicotômico de espécies reservatório ou não foi sendo desafiado durante o século passado, porquê havia variação na resposta dos hospedeiros aos parasitos, em dependência a fatores já discutidos neste livro: ambiente, comunidade parasitária, organização social e alimentação dos animais hospedeiros. Havia um sucesso ecológico diferente de parasitas e hospedeiros que não podia ser estimado adequadamente baseado no modelo de animais reservatórios naturais, mas sim em Sistemas Reservatório: uma associação de diferentes espécies de hospedeiro, cada uma delas com diferentes suscetibilidades a uma determinada comunidade parasitária (Ashford 1997; Haydon et al. 2002).
Nós podemos mensurar o sucesso ecológico de um determinado organismo em um dado ambiente por variáveis que modificam de forma relevante a pirâmide de vida (vida Cap) de uma população biológica: a Natalidade e a Mortalidade de tal população, tanto gerais, como contingenciadas pela idade ou sexo dos componentes da população. Através do estabelecimento de uma razão entre a taxa de natalidade e a de mortalidade de uma determinada população biológica (ambas medidas na mesma escala de tempo, evidentemente) nós podemos estimar se esta população está crescendo ou diminuindo, através do cálculo da Taxa Reprodutiva Básica (J. A. P. Heesterbeek 2002), como demonstrado na forma da Equação (6.1):
R∅=Tx FertilidadeTx Mortalidade
A Taxa Reprodutiva Básica é conhecida como R∅ (ou R naught) (J. a. P. Heesterbeek and Dietz 1996), e podemos adaptá-la a diferentes contextos, dentro do mesmo conceito: de que uma população biológica está crescendo se a taxa de nascimentos é superior à de mortes. A epidemiologia utiliza a Taxa Reprodutiva Básica conforme a definição abaixo (Delamater et al. 2019):
“R∅ é o número médio esperado de casos secundários produzidos por um hospedeiro infectado típico, durante todo seu período infeccioso, dentro de uma população completamente suscetível (J. a. P. Heesterbeek and Dietz 1996).”
A ideia por trás deste conceito é que uma infeccção estará estável na população se a partir de cada hospedeiro infeccioso é produzido exatamente uma nova infecção. O valor de R∅ desta infecção seria exatamente 1 (um) no período considerado. Equivale a dizer também, por um viés epidemiológico, que a taxa de produção de novas formas infectantes do parasito produzidas por unidade de tempo equivale ao número de formas infectantes destruídas no mesmo espaço de tempo. Se a taxa de produção é igual à taxa de destruição, a população permanece estável. Já a infecção estará crescendo na população infectada se a partir de cada hospedeiro infeccioso é produzido mais de um hospedeiro infectado. Neste caso o valor de R∅ será maior do que 1 (um). Quanto maior o valor de R∅ acima de um, mais rápida é a progressão da infecção na população afetada, porquê haverá maior número de novos infectados por hospedeiro infectado, por unidade de tempo. Por outro lado, quando o valor de R∅ for menor do que 1 (um), significa que a proporção de infectados na população se reduzirá gradativamente, gerando tanto menos novos infectados a partir de cada animal infeccioso quanto mais próximo de zero for o valor de R∅. Finalmente, quando o valor de R∅ for equivalente a zero, nenhum novo hospedeiro infectado é produzido a partir de um hospedeiro infeccioso. Isto equivale à dizer que a infecção tende a desaparecer no curso do tempo, por inexistência de novos animais infecciosos.
Note que O valor de R∅ do parasito é pois uma resultante de uma plétora de fatores que intermediam a relação hospedeiro - comunidade parasitária albergada - população parasitária específica, e ele reflete o sucesso ecológico da espécie de parasito em questão. Esta resultante será diferente para cada população parasitária infectando cada população hospedeira, com o valor de R∅ para cada espécie de parasito sobre uma população hospedeira sendo a média dos valores de R∅ para cada hospedeiro infectado individualmente. Em um ambiente natural diverso esta variação de R∅s torna determinadas espécies de hospedeiros mais competentes na manutenção e dispersão do parasito (R∅s acima de 1), enquanto outras não serão tão competentes nestes quesitos (R∅ <1). Algumas espécies serão completamente incompetentes para manter ou dispersar o parasito (R∅ = 0). A reunião conjunta no ambiente destas diversas competências de transmissão passou a ser compreendida pela Epidemiologia Moderna como um “Sistema Reservatório”, onde cada hospedeiro contribui de forma diferencial para a manutenção e dispersão do parasito no ambiente (Ashford 1997; Haydon et al. 2002), baseado em seu valor de R∅ para o parasito, conforme a Tabela 6.1. Neste “Sistema Reservatório” hospedeiros “Reservatório” mantém e dispersam o parasito no ambiente, mas não são os únicos hospedeiros lá existentes. Hospedeiros de “Espalhamento” não amplificam a infecção ou a contaminação do ambiente, mas dispersam o parasito carreando-o e permitindo o contato com novos hospedeiros suscetíveis. Hospedeiros ditos “Finais” são incapazes de manter ou dispersar o parasito atenuando a infecção no ambiente, por não serem oportunidades de reprodução para o parasito.
| R∅ | Situação da Infecção | Tipo de Hospedeiro |
|---|---|---|
| Maior do que 1 | Crescendo | Reservatório |
| Igual a 1 | Estável | Espalhamento |
| Entre 0 e 1 | Diminuindo | Espalhamento |
| Igual a 0 | Finalizada | Final |
| a Valores de R∅ decimais devem ser utilizados como proporções: um valor de 1,1 equivale a dizer que a cada 10 animais infecciosos, onze novos animais serão infectados, ou 110% da população original. Um valor de 0,5 significa que a partir de 10 animais infecciosos apenas 5 animais seriam novos infectados, ou 50% da população infecciosa original. |
Estas diferenças de R∅ entre cada hospedeiro individual geralmente apresentam uma distribuição populacional desviada em relação à média. Isto significa que em uma população hospedeira, uma pequena proporção dos indivíduos apresenta altos valores de R∅, enquanto o restante apresenta valores muito menores (Poulin 2004; McVinish and Lester 2020). Há uma agregação na produção de formas infectantes do parasito em uma pequena parcela da população. Este fato é relevante para estratégias de controle parasitárias: a Medicina Veterinária moderna precisa investir na identificação, tratamento ou remoção destes indivíduos hospedeiros que são grandes dispersores do parasito localmente, do que tratar indistintamente todo o plantel sob vigilância.
Esta teia de hospedeiros transmissores com diferentes competências de uma grande diversidade de parasitos dá origem também a “Teoria do Fator de Diluição Ambiental”: quanto mais diverso um ambiente hospedeiro mais difícil é a invasão de novos parasitos e o estabelecimento de epidemias, porquê a diversidade de espécies “tamponaria” (atenuaria) o potencial epidêmico de cada parasito individualmente (Schmidt and Ostfeld 2001; LoGiudice et al. 2003; F. Keesing, Holt, and Ostfeld 2006; Felicia Keesing and Ostfeld 2021). Em última instância, como o que nos interessa é o hospedeiro, é este que precisa ter sucesso ecológico frente à comunidade parasitária que o infecta, desafia ou colabora, para que seu R∅ seja maior do que 1 e sua população progrida ecologicamente.
6.2 Quando a doença ocorre?
A distribuição da ocorrência da doença no tempo pode nos fornecer muitas informações relevantes acerca do processo mórbido em análise. Entenda que “distribuição no tempo” é uma referência que pode ser variável, dependendo da escala de tempo que o pesquisador escolher, ou que se mostrar mais apropriada no caso em questão. O que o pesquisador deve procurar é agregações dos casos da doença no tempo. Estas agregações podem estar associadas a escalas de tempo diferentes:
Diárias: os casos da doença concentram-se em determinados dias ou horas do dia. Um exemplo clássico deste tipo de agregação, ou sincronia (Greischar, Read, and Bjørnstad 2014), é a Malária. Dependendo da espécie de Plasmodium que estiver afetando o animal, há uma concentração dos sinais clínicos em ciclos de 2 dias (P. falciparum, P. vivax, and P. ovale) ou de 3 dias (P. malariae), que coincidem com a lise das hemácias causada pelo hematozoário. Veja que estes fenômenos são típicos de seleção natural, não envolvendo nenhuma forma de “querer” do Plasmodium, mosquito ou hospedeiro. Serão favorecidos os clones do hematozoário que lisarem as hemácias na hora em que o mosquito está atuando mais intensamente, sendo naturalmente removidos do pool genético os clones que lisarem as células fora de hora, por não alcançarem o vetor invertebrado.
Sazonal: Muitas doenças demostram uma concentração em determinadas épocas do ano, devido à influência dos fatores abióticos que geram diferenças sazonais. Ambientes com diferenças bem demarcadas de temperatura (inverno/verão), pluviosidade (estação seca/chuvosa), ou mesmo vento intenso (presente/ausente), podem gerar ciclos de ocorrência de doença que se alternam entre tais categorias. A temperatura menor no inverno acarreta um aumento das doenças respiratórias, tanto por aumentar a sobrevivência de formas infectantes de parasitos, como por aumentar a demanda energética para geração de calor corporal, reduzindo a quantidade de energia calórica que pode ser direcionada para o sistema imune, ou mesmo porquê os animais agloram-se mais, aumentando a transmissibilidade dos parasitos entre indivíduos. Já no outro extremo, a estação quente (que muitas vezes é também chuvosa) aumenta o metabolismo dos invertebrados em geral e todas as doenças em que eles contribuem com a ocorrência, seja como agentes diretos (como a hematofagia causada por carrapatos, pulgas) ou indiretos (onde podem atuar como vetores mecânicos ou hospedeiros intermediários de doenças). Locais com uma estação chuvosa bem demarcada tem aumento das doenças transmitidas por hospedeiros invertebrados nestas épocas, como Malária, A Leptospirose é uma doença que também apresenta um aumento de incidência nas épocas chuvosas, mas devido aos alagamentos e consequentes espalhamento das Leptospiras contidas na urina dos roedores para o alimento e ambientes frequentados pelos animais que serão eventualmente afetados. O ambiente mais encharcado também pode aumentar a frequência de doenças dos membros locomotores (que ficam encharcados ou enlameados durante muito tempo), digestórias (devido ao apodrecimento do alimento, contaminação de pasto, ou mesmo mudanças de composição ou contaminação da água de bebida). Locais com alta intensidade de vento em determinadas épocas, como o Minuano no inverno do Sul do Brasil (ou o Pampero na Argentina), o Sirocco na Espanha, o Mistral na França, todos são exemplos de ventos fortes, constantes e sazonais que podem gerar doenças devido ao frio ou calor intenso, desidratação, ou mesmo pelo carreamenteo de formas infectantes de parasitos para onde o vento sopra.
Cíclicas: Tais agregações de doenças geralmente são associadas a eventos ambientes graves ou intensos, mas sem periodicidade demarcada. Doenças podem demonstrar agregação após grandes tempestades, ondas de frio ou calor, eventos anômalos de produção humana, como derramamentos de petróleo, gases tóxicos ou radiação. Em todos estes casos, os animais afetados vão demonstrar os sinais clínicos resultantes da alteração ambiental de forma concentrada em um determinado intervalo de tempo após a alteração. O pesquisador deve estar atento para poder relacionar tais eventos aos quadros mórbidos que ele eventualmente esteja estudando.
6.3 Onde a doença ocorre?
A localização geográfica da doença na população de hospedeiros afetados sempre foi considerado pelos epidemiologistas, desde Hipócrates (águas, ares e LUGARES), passando por John Snow (que referenciou no mapa de Londres a localização dos casos de cólera humana e os relacionou à localização das fontes de água), como fator preponderante para explicar a ocorrência da doença. Esta importância do estabelecimento da localização geográfica da doença em questão ganhou importância adicional a partir da Teoria da Higiene ser estabelecida, já que o agente etiológico, o causador estaria associado a determinadas condições ou situações de ambiente. A partir da criação e estabelecimento das ferramenteas moleculares a Teoria da Nidalidade pressupõe que a partir de uma modificação molecular, genética, do microorganismo é que pode se estabelecer a doença ou variante da doença mais patogênica ou de maior transmissibilidade. Assim, o Nidus, o local de surgimento da doença novamente vê sua importância ressaltada. Nossa geração de Epidemiologistas tem agora à disposição ferramentas poderosas de análise geoespacial, onde aplicativos de computador são capazes tanto modificar e filtrar imagens de satélite (tanto em tempo real, como séries históricas) como de analisar e modelar estatisticamente tais imagens de monitoramento via satélite, além da própria facilidade de georeferenciamento do hospedeiro ou agente parasitário de interesse. Esta união deu origem a praticamente um ramo novo da Epidemiologia, a Análise e Modelagem Geoespacial das doenças.
Para o fim de análise geoespacial a informação mais importante é a geolocalização dos hospedeiros afetados. A partir do estabelecimento da rede mundial de geoposicionamento por satélite (GPS) na década de 80 do século passado, uma rede de satélites que iniciou-se com fins militares, mas que gradualmente adquiriu importância preponderante para vários processos tecnológicos da espécie humana que dependem de navegação ou localização precisa, geolocalizar qualquer ponto no globo terrestre ficou extremamente fácil. Plantadeiras e colheitadeiras agrícolas navegam hoje em dia orientadas por sistemas GPS; nenhuma construção moderna de maior porte é feita sem uma definição preecisa de seus marcos por GPS. Qualquer celular de mão possui um sistema GPS incorporado, e aparelhos GPS mais sofisticados podem ser adquiridos a custo muito baixo. Assim, geolocalizar o ponto geográfico preciso onde o animal morreu, saber por onde ele se deslocou e/ou o tempo gasto em cada local geográfico (a partir de séries de geolocalizações), mesmo em tempo real (através de coleiras equipadas com GPS), são informações facilmente obtidos em nosso século. Podemos mesmo estimar a velocidade de deslocamento de determinado animal, e estimar quando ele ficou doente a partir de uma mudança de padrão ou velocidade de deslocamento se sob a influência de uma determinada doença.
Geolocalizar um ponto específico de interesse pode ser feito tanto de forma direta como indireta. A forma direta exige que o pesquisador esteja ou possa dirigir-se pessoalmente ao ponto em que o animal está doente ou morto. No local ele fará a leitura (com auxílio de equipamento adequado que, como dissemos, pode ser mesmo seu próprio aparelho celular) das coordenadas geográficas onde está, anotando-as em papel, enviando-as para a “nuvem de informações” online, ou registrando-as em software adequado. Esta geolocalização direta é vantajosa para o pequisador porquê permite a ele examinar diretamente o animal afetado/morto, fotografá-lo e ao ambiente circundante in loco, eventualmente coletar material biológico, entrevistar tratadores/proprietários/tutores/moradores locais sobre as características da doença nos animais afetados, coletar amostras ambientais de água, solo, excretas, plantas, reunindo pessoalmente um cabedal de informações preciosas para chegar ao diagnóstico final do que pode ter levado aquele animal à morte, ou do quê pode estar afetando aquela população de animais em particular.
Já a geolocalização indireta é realizada a partir de uma informação transmitida ao pesquisador de aonde o animal ou população está. O pesquisador por sua vez atribui uma coordenada geográfica correspondente a esta informação que a ele foi comunicada. Esta informação transmitida pode ter um grau de incerteza variável. Informações sobre animais mortos ou doentes em cidades ou povoações humanas geralmente vem acompanhadas de algum tipo de endereço ou referência que permite uma geolocalização indireta relativamente precisa. Já animais encontrados em situações mais rurais ou em ambientes naturais podem tem graus de incerteza muito grandes, e a coordenada geográfica atribuída a tal informação pode estar a quilômetros de distância do ponto real de encontro do animal.
Uma forma de atenuar tais discrepâncias na análise final é atribuir a localização do animal não a um ponto geográfico determinado, mas a uma outra unidade de organização geográfica que seja representativa de onde o animal morto ou doente foi visto. Assim, quarteirão, rua (ou trecho de rua), bairro, município, propriedade ou parte de propriedade, todos são unidades geográficas a que podem ser atribuídas o local de encontro do animal. A comparação entre os locais de encontro dos animais tem que ser feita pela unidade geográfica de menor precisão entre as que foram utilizadas. Por exemplo, se eu localizei entre dez animais mortos, cinco por coordenadas geográficas e cinco por rua, todas as comparações devem ser feitas levando a rua em consideração. Se cinco foram localizados por barracão de propriedade e cinco foram por município, o município é que deve ser a unidade de comparação geográfica. É claro que há uma perda de precisão geográfica em todos estes casos. O pesquisador deve julgar se tal perda é razoável para a pergunta que seu trabalho de pesquisa pretende responder. Digamos que no caso de em dez animais, nove foram geolocalizados precisamente por coordenadas geográficas e um por município, provavelmente deve valer a pena descartar na análise esta única informação de menor precisão para preservar a qualidade das outras nove que a tem em alto grau.
Note-e que quando há esta ampliação do georeferenciamento do ponto de ocorrência da doença, informação pode ser perdida. Digamos que uma determinadad doença esteja associada à proximidade a cursos ou coleções d´água, situação típica da Leptospirose, p.ex. Se eu georeferenciar os casos de Leptospirose por coordenada geográfica precisa, será fácil associar a proximidade da água à doença. Mas se eu utilizar um critério de georeferenciamento por uma escala mais ampla, esta informação pode ser diluída no ambiente englobado pela escala utilizada. Dentro de um bairro, dentro de uma rua, no caso destas forem as unidades utilizadas, podem haver diferentes proximidades a locais úmidos, impedindo o relacionamento da doença (no caso a Leptospirose) a esta característica.
Que fique claro que a grande importância da geolocalização da doença ou do animal doente não é a coordenada geográfica per se mas sim quais são as condições das variáveis abióticas e bióticas naquele ponto em questão. São altitude, vento, clima, constituição do solo, insolação, vegetação, fauna e microbiota local, entre qualquer variável que o pesquisador venha a considerar relevante localmente que nos interessam e não meramente a Latitude e Longitude do local. Assim, pode extrair tais informações tanto localmente, quando da possibilidade de presença no local do georeferenciamento, quanto digital ou remotamente através de imagens de satélites, mapas, atlas, catálogos, bancos de dados de pesquisa ou empresariais é que será o desafio do Epizootiologista aplicado.
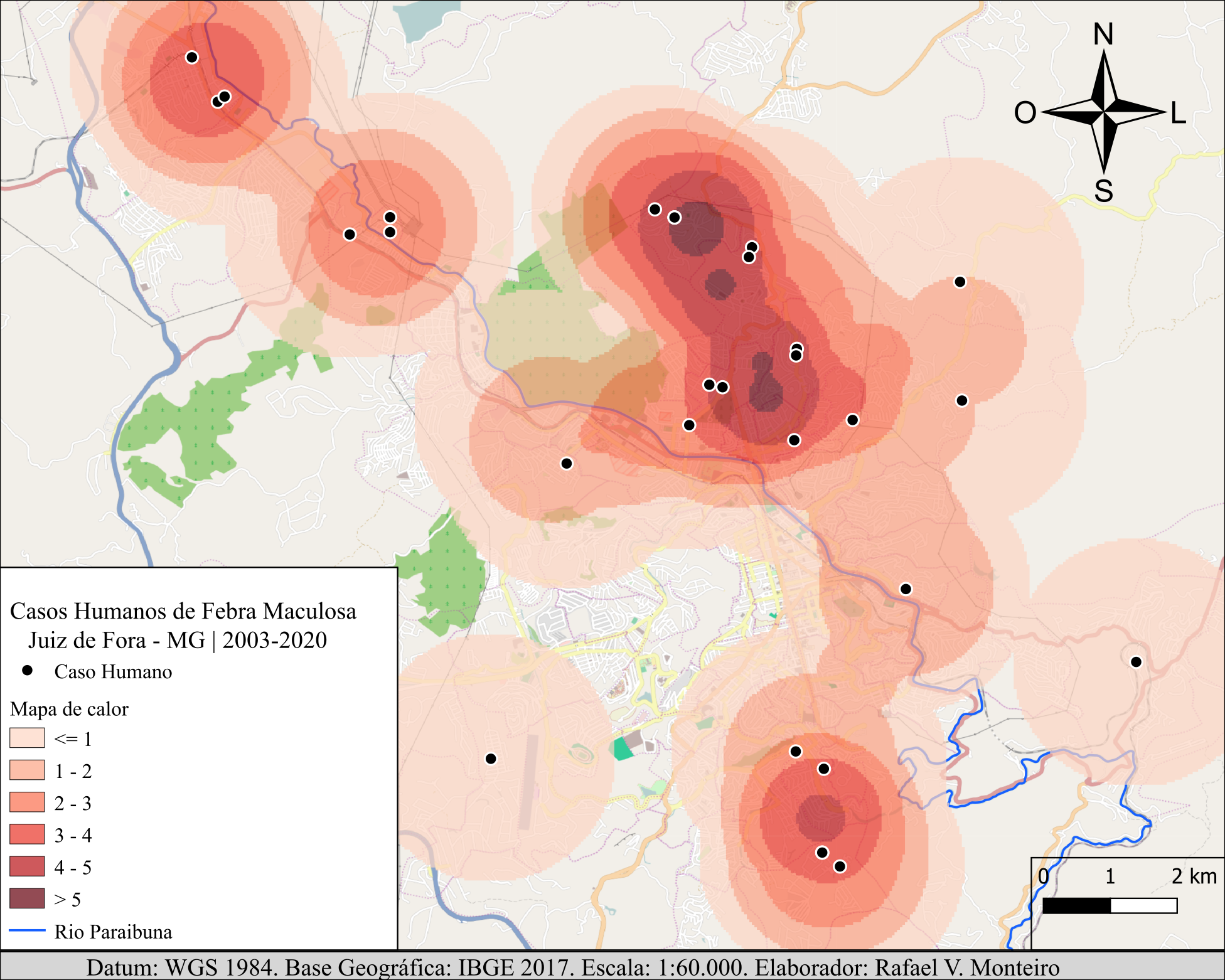
Figura 6.1: Distribuição geoespacial histórica (2003-2020) de casos de Febre Maculosa humana no município de Juiz de Fora (MG). Os casos demonstraram um padrão de agregação no espaço (análise do vizinho mais próximo com padrão agregado) e no tempo (mapa de calor de sobreposição de ocorrência histórica). Tal tipo de análise pode revelar características geoespaciais relevantes para a ocorrência da doença, como foi o presente caso. Mapa redesenhado a partir dos mesmos dados de Rafael Veríssimo Monteiro et al. (2022).
Monteiro e colegas (Rafael Veríssimo Monteiro et al. 2022) utilizaram esta abordagem para demonstrar que os casos humanos de Febre Maculosa no município de Juiz de Fora (MG) apresentavam um padrão agrupado no tempo e no espaço, revelado pela análise do vizinho mais próximo (NNN - Nearest Neighbour Analysis; Figura 6.1). Este resultado sugere a existência de fatores epidemiológicos, associados a estas regiões mais “quentes” (hotspots), que aumentam o risco para a ocorrência de casos de Febre Maculosa humana. No caso em questão, tais regiões possuiam IDH mais baixo, e as baixas condições de higiene e moradia representavam um fator de risco para a exposição aos carrapatos infectados. Estes dados também sugeriram que, no cenário local, os cavalos domésticos são mais relevantes como reservatórios da bactéria causadora da doença do que as capivaras residentes nos cursos d´água locais.
6.4 Como e Porquê a doença ocorre
Por tais duas perguntas o Epizootiologista deve entender a patogenia da doença. Em nosso século respondê-las tornou-se muito mais fácil do que nos anteriores, tendo em vista a miríade de diferentes ferramentas de análise de funções orgânicas que possuímos atualmente. Entretanto, uma dificuldade continua presente, que não mudou muito com nossas ferramentas atuais: a necessidade de amostras biológicas de qualidade. Na prática veterinária um pesquisador pode ter acesso a um animal vivo ou morto para realizar um diagnóstico. Discutiremos aqui o diagnóstico nas duas situações.
6.4.1 Diagnóstico pré-morten
O diagnóstico realizado com o animal ainda vivo pode ser feito de forma direta ou indireta. A abordagem direta pressupõe a coleta de material biológico diretamente do animal doente. Esta é normalmente a abordagem tradicional da Medicina Veterinária para diagnóstico de enfermidades, e apenas falarei aqui em linhas gerais sobre tal procedimento e ações associadas, visto que este não é o objetivo principal deste livro e sobre o que já que existem inúmeros livros e artigos científicos versando especificamente sobre o diagnóstico de enfermidades. Alguns critérios devem ser respeitados para a coleta do material biológico:
- Que tal material seja representativo do sistema orgânico que está comprometido no animal, ou seja que ele seja capaz de carrear consigo indicativos da doença ou do agente etiológico.
- Que a quantidade do material coletado seja suficiente para permitir a detecção de alteração ou da presença do microorganismo ou característica da doença, ou seja que o limite mínimo de detecção do teste utilizado seja atingido.
- Que ele seja conservado adequadamente até análise, ou seja que o material coletado seja mantido em temperatura ou técnica de conservação que preserve as características da doença ou do agente etiológico que deseje-se detectar.
- Que o tempo de conservação não ultrapasse a degradação do material ou dos indicadores da doença, que mesmo sob conservação ocorrem naturalmente.
Normalmente, sangue é o material biológico “universal”, já que é um tecido orgânico que entra em contato com todos os outros tecidos do organismo, carreando consigo microorganismos (príons, vírus, bactérias, fungos e protozoários) e/ou seu material genético, proteínas e/ou peptídeos, células (alteradas pela doença ou não), além de gases e íons, em suma, vários indicativos de doença ou morbidade oriundos dos tecidos e sistemas orgânicos alterados. Biópsias tegumentares e punções seguidas de biópsia transcutâneas são utilizadas para a coleta de pele, tecido muscular e hepático, exsudações e fluidos articulares e abdominais. Outros tipos de biópsias (geralmente dos sistemas cárdio-respiratório, digestório, renal, reprodutivo, neurológico, hormonal, articulares) já exigem anestesia geral e analgesia, por envolverem normalmente o acesso à cavidade abdominal, caixa craniana ou às articulações. Finalmente, fezes e urina são dois materiais excretados pelos animais que podem fornecer informações de qualidade para o diagnóstico de enfermidades. Entretanto, para a coleta de sangue ou outros materiais biológicos é necessária a “colaboração” do animal, que, no caso de animais domésticos é facilmente obtida com algum carinho e contenção mecânica. Já no caso de animais silvestres, ferais, de criação extensiva ou de grande tamanho, tal cooperação pode ser difícil, exigindo contenção mecânica mais rigorosa ou mesmo contenção química (tranquilização ou anestesia geral). Em todos os casos anteriores, a coleta já se torna mais difícil de ser realizada, ou mesmo impossível, dependendo do animal, condições de manejo, disponibilidade de drogas, equipamentos e instalações. Mesmo o sangue coletado nas condições descritas pode sofrer alterações devido à reações adrenérgicas agudas de estresse ou resultantes do uso dos anestésicos. A abordagem indireta coleta, do ambiente frequentado pelo animal ou das instalações que ele utiliza, fluidos, secreções e excreções que possam ser obtidas, de forma que estas possam fornecer informações sobre o tipo de agente parasitário ou de afecção está incidindo sobre o animal analisado. Note-se que a abordagem indireta, dependendo do número de animais que frequente o local da coleta ambiental, pode não ser exclusivo ou mesmo representativo do animal doente, fato que deve ser levado em consideração na interpretação dos resultados obtidos.
6.4.2 Diagnóstico pós-morten
No caso do animal já morto, o tempo decorrido desde a morte e o estado de conservação da carcaça são duas características que interferem sobremaneira na capacidade do patologista dar respostas acuradas sobre as alterações mórbidas que o animal possuía antes de morrer. A partir do momento em que cessam batimento cardíaco e respiração as células do organismo começam a alterar-se por causas tanatológicas e não causas ligadas à doença que o animal apresentava antes de morrer. Ademais, toda a microbiota residente em tal organismo, livres dos controles fisiológicos, imunológicos, mecânicos, entre outros, impostos pelo organismo que as albergava, começam a provocar alterações nos tecidos que também são tanatológicas e não mórbidas. A diferenciação entre alterações pré-morten e pós-morten tornam-se gradualmente mais difíceis de serem realizadas por técnicas histológicas, citológicas, moleculares, entre as várias que podem ser utilizadas no estudo de uma carcaça, até o ponto em que as alterações pós-morten são predominantes e impedem qualquer tentativa de diagnóstico do que o animal tinha quando em vida. É claro que determinados processos mórbidos, principalmente os traumáticos, deixam marcas indeléveis mesmo na carcaça mais degradada. Ossos são sempre as testemunhas finais da vida de um vertebrado, e fraturas, lesões por projéteis de arma de fogo, alterações típicas de determinadas doenças metabólicas ou infecciosas que deixem marcas ósseas podem ser caracterizadas em fósseis e restos mumificados de animais. Infelizmente estes são casos raros, mas que ainda assim podem contribuir para o diagnóstico de um quadro mórbido em um animal.
A conservação adequada da carcaça deve ser portanto preocupação preponderante do Epizootiologista. A situação ideal é a necrópsia imediata do animal recém morto. Esta situação ideal infelizmente não é comum. Vários fatores contribuem negativamente para impedir esta atuação ideal. Podemos elencar alguns deles. Atente-se ao fato de que a capacidade de encontrar, conservar, necropsiar e coletar material biológico de qualidade de um animal é função de vários deles, e não de apenas um só:
Local de residência ou alojamento do animal: animais internados em instalações hospitalares, domésticos, de cativeiro e de produção intensiva são monitorados muito mais intensamente do que animais silvestres, caipiras ou de vida livre. Nos primeiros é muito mais provável que alguém detecte a morte do animal e o encaminhe prontamente para a necrópsia ou que tome alguma atitude para melhorar a conservação da carcaça (resfriamento, congelamento, administração de soluções conservantes) até que a necrópsia possa ser realizada. No caso dos outros grupos de animais a rapidez de encontro do animal pode ser muito baixa, comprometendo a conservação da carcaça.
Tamanho do animal: Animais de pequeno porte degradam-se tremendamente mais rápido do que animais de porte maior. A pequena massa corporal de pequenas aves, répteis, peixes e mamíferos acarretam uma rápida dessecação, destruição por insetos (formigas e outros podem destruir um pequeno passarinho durante a noite), predação por animais carniceiros, além destas pequenas carcaças sofrerem alteração pós-morten pela invasão da microbiota própria em poucas horas. Já em animais de porte maior todos estes efeitos são atenuados pela maior massa corporal, pele mais grossa, tamanho dos órgãos maiores, e assim por diante. Tamanho também interfere no próprio ato de encontrar o animal. Animais pequenos podem ser difíceis de encontrar mesmo em ambientes pequenos e controlados, dependendo da ornamentação ou complexidade do local. Já animais grandes são naturalmente mais fáceis de serem encontrados, mas mesmo animais de grande porte como grandes herbívoros, felinos e outros grandes taxa podem passar despercebidos em situações naturais.
Temperatura ambiental: Animais que venham a morrer em ambientes frios ou com temperatura sub-zero podem ser naturalmente conservados, principalmente no segundo caso, pelo congelamento rápido que será imposto à carcaça. Já nas regiões tropicais, semi-áridas e áridas o inverso ocorre: rápida degradação da carcaça por temperaturas altas, dessecação ou umidade excessiva, rápida invasão pela microbiota autóctone dos tecidos orgânicos, predação por invertebrados e vertebrados, carreamento e/ou degradação por chuvas intensas. Veja que estes efeitos impostos por temperaturas e umidades mais altas podem acontecer mesmo com animais alojados em ambientes controlados.
Treinamento do profissional para ver o animal: Neste item a experiência é fundamental. Um profissional com nenhum ou pouco treinamento vê animais muito menos do que um profissional com larga experiência. Esta é uma questão intrínseca do treinamento do próprio SNC para reconhecer padrões de cor, forma, tamanho como relativos a determinados animais, e é impossível antecipar no profissional sem experiência quantos animais ele conseguirá ver, se algum verá.
Além do próprio encontro do animal os fatores acima podem impedir a conservação da carcaça por problemas indiretos: é fácil andar com uma caixa térmica em qualquer situação e ir colocando pequenos vertebrados dentro de sacos e na caixa. Mas o que fazer quando um boi, um grande cachorro, um veado é encontrado? A caixa torna-se inútil, a não ser para conservação de fragmentos de órgões. A necropsia à campo é a solução para estas situações em que o tamanho do animal, a distância e condições de locomoção até a instalação de pesquisa, ou mesmo o risco biológico de transportar o animal até outro lugar impediriam uma necrópsia adequadamente realizada em um laboratório ou instalação médico-veterinária. A necropsia a campo auxilia tremendamente no diagnóstico, já que com uma caixa contendos EPIs, instrumentos cirúrgicos, material de desinfecção, um bom conjunto de facas e instrumentos de afiação, frascos de conservação de diferentes tamanhos e líquidos conservantes adequados (como soluções de formoldeídeo, glutaraldeídeo e outras mais especializadas para conservação de material genético), um profissional treinado pode coletar material biológico de alta qualidade e transportá-lo perfeitamente conservado até o laboratório.
6.5 Quais os parasitos envolvidos?
A evolução da compreensão da parasitologia e da infectologia sobre os processos orgânicos que acarretam as doenças infecciosas ressaltam cada vez mais a importância da comunidade parasitária que infecta algum animal sobre a ocorrêncida da doença nele. A ideia de “um parasita-uma doença” é uma ideia obsoleta frente à esta compreensão mais avançada de que uma séria de interações de cooperação e competição ocorrem entre os parasitas e destes com seu hospedeiro, e que a doença é uma resultante de tais interações. O desafio do epizootiologista moderno é deslindar tais interações, encontrando pontos de interferência em tais comunidades que sejam positivos para a saúde do hospedeiro, e não meramente a tentativa de eliminar determinado parasito.
Mas para chegarmos a este ponto de compreensão da teia de interações causais que podem estar afetando algum hospedeiro ou populações de hospedeiro, nós precisamos identificar os parasitos, para estimar a intensidade de seus efeitos sobre a saúde daqueles, e isto pode ser feito através de algumas técnicas. Nossa evolução na identificação parasitária foi grande nos últimos 300 anos. As primeiras técnicas parasitológicas eram dependentes apenas da capacidade de visualização humana, limitadas destarte por nossa acuidade visual. O olho humano é capaz de distinguir organismos com pelo menos 100 micrômetros; tipicamente pequenos detalhes em mapas, por exemplo, tem 300 micrômetros. Assim, nosso estudo dos organismos que infectavam humanos e animais e que podiam ser associados à doenças limitavam-se a pequenos ácaros, piolhos, pulgas, ou seja, artrópodes em geral.
A partir da evolução da óptica e da criação das primeiras lentes de aumento, culminando na invenção do microscópio óptico, nós pudemos identificar, distinguir, descrever e caracterizar um grande número de novos agentes parasitários. Duas novas áreas da ciência foram criadas e evoluiram: a microscopia óptica e a microbiologia. Estes ganhos de visualização óptica somados à teoria da higiene permitiram a associação de tais parasitos à doenças, inclusive com a caracterização microscópica. A identificação das características microscópicas de protozoários, fungos e bactérias, em muito suportaram a infectologia no início do século XX. Mas mesmo o microscópio óptico tem seus limites de discriminação visual, devido às características de lentes, distorções visuais, artefatos ópticos, e emissão do feixe luminoso. Tipicamente um microscópio óptico consegue discriminar detalhes de no máximo 500 nanômetros.
Entretanto, mesmo esta grande capacidade de discriminação óptica não conseguia caracaterizar vírus nem tampouco príons. As suspeitas de uma geração de microbiologista sobre parasitos muito pequenos para identificação microscópica, mas que deixavam seus sinais citopáticos e tinham doenças associadas. A confirmação dos vírus como os parasitos que eram procurados só pode ser atingida através da microscopia eletrônica, já em meados do século XX. Esta técnica evoluiu tremendamente deste então, e já existem equipamentos e software associado que permitem a discriminação de detalhes dos núcleos dos átomas e suas elestrosferas. Assim, a capacidade atual de discriminação de agentes causadores de doença vão até um tamanho atômico, se tal agente existisse. O último grupo de parasitos, se assim podem ser chamados, a serem descobertos são os príons. Tais proteínas causadoras de deonça são proteínas modificadas auto-replicantes ou que polimerizam proteínas normais transformando-as em modificadas. Esta modificação é associada à formação de placmas mielóides nos neurônios dos SNC, sua acumulação intra- e peri neuronal, culminando com a morte destes neurôniocs e vacuolização do SNC. As Encefalites Espongiformes já foram caracterizadas em várias espécies animais, sempre associadas ao processo acima descrito: mutação de proteína cerebral (adquirida ou expôntanea), expansão da mutação nos tecidos do SNC e vacuolização do encéfalo.
A visualização dos parasitos normalmente é feita nos tecidos orgânicos examinados por técnicas histológicas, como esfregaços sanguíneos, cortes teciduais montados em lâminas de microscopia, ou cortes teciduais preparados para a microscopia eletrônica. A determinação de índices de presença ou intensidade de ação parasitária podem ser criados a partir da contagem de quantos parasitos eu visualizo ou estimo presença, dividido por um denominador adequado, como número de células analisadas, área ou volume de tecido analisado. Tais índices parasitários permitem a estimativa de que, sobre determinada espécie/taxa animal, haja uma maior citopatogenicidade e/ou produção de novas formas infectantes do parasito em determinado órgão, local ou tecido, logo causando doença. Os mesmos índices podem ser usados comparativamente entre parasitos diferentes, de forma a estimar a severidade ou letalidade de determinada infecção. O contingenciamento dos resultados por categorias de hospedeiro, como sexo, idade, ou por modificadores, como co-infecções parasitárias, deve sempre que possível realizado, já que a moderna epidemiologia prevê a presença e importância de tais interações.
Além das técnicas de visualização direta para confirmação de infecção, podemos diagnosticar a presença de parasitos através da caracterização dos efeitos teciduais e clínicos sobre o organismo do hospedeiro ou dos sinais moleculares da presença parasítica atual ou passada. A descoberta da técnica da reação de polimerase em cadeia ou PCR, e o posterior desenvolvimento das técnicas de sequenciamento de gens e genomas, revolucionou o estudo dos parasitos. Agora o pesquisador pode identificar um determinado agente parasitário com crescente facilidade, sensibilidade e acurácia, através de identificação de DNA/RNA do parasito, seguido do sequenciamento de diferentes proporções de tal material genético, indo desde um simples gene até genomas inteiros. A técnica de PCR é frequentemente muito sensível, sendo capaz de identificar a presença de um único parasita em uma amostra relativamente grande de tecido.
A última estratégia técnica utilizada para diagnosticar parasitas são as técnicas que detectam anticorpos, em soro ou plasma geralmente, indicativos de que o organismo estudados é portador ou teve contato com determinado parasito. Tais técnicas tem graus de especificidade de diagnóstico parasitário e de sensibilidade na identificação de anticorpos muito variáveis e podem apresentar resultados discrepantes entre indivíduos ou tipo de teste mais frequentemente. Repetições conferem sempre mais solidez aos resultados, como uma forma de atenuação de fontes de variabilidade espúria nas análises realizadas. É claro que com dados obtidos por técnicas correlacionais podemos sugerir apenas a grande coincidência entre causa (parasito identificado/contado no tecido) com o efeito (lesão nas células do tecido com o parasito). Esta é a qualidade de informação obtida com as técnicas epidemiológicas transversais: correlação na frequência de ocorrência. Para podermos estabelecer uma associação causal entre parasito (ou associação de parasitos) e doença são necessárias técnicas epidemiológicas que incluem a identificação individual do hospedeiro, e seu acompanhamento no curso do tempo. Assim é estabelecida a situação clínica e bioquímica dos tecidos de um animal antes da entrada em contato com determinado parasito e verifica-se que elas foram modificadas negativamente devido a efeitos da atuação do parasito após sua entrada no organismo. Só assim podemos confirmar a existência de uma relação causal inequívoca entre o início da presença do parasito no hospedeiro e a consequente lesão tecidual que passe a existir.