Topic 5 Protein Translation: from RNA to Proteins
5.1 Pre-Lecture Concepts
5.1.1 Aminoacyl-tRNA Synthetase
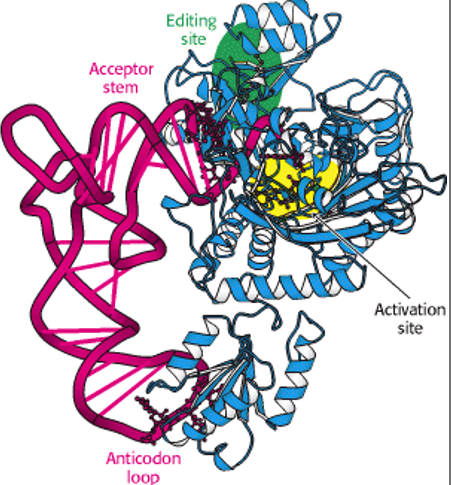
Figure 5.1: Aminoacyl-tRNA Synthetases
Aminoacyl-tRNA synthetases are a group of enzymes that attach amino acids to tRNAs. Each tRNA molecule is recognized by a specific aminoacyl-tRNA synthetase, each of which can aminoacetylate different tRNAs whose anticodons correspond to the same amino acid.
For this reason, the chemical reaction that leads to amino-acetylation occurs in two steps:
An activated amino acid intermediate is formed by reaction between amino acid and ATP
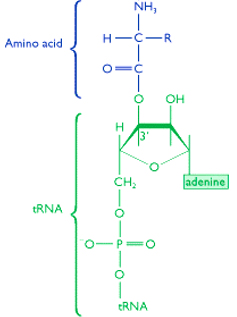
The amino acid is then transferred to the 3’ end of the tRNA
Note that the link is formed by the COOH group of the amino acid and the OH group that is attached to either the second or the third carbon on the sugar of the last nucleotide (which is always an “A”).
5.1.2 Ribosomes
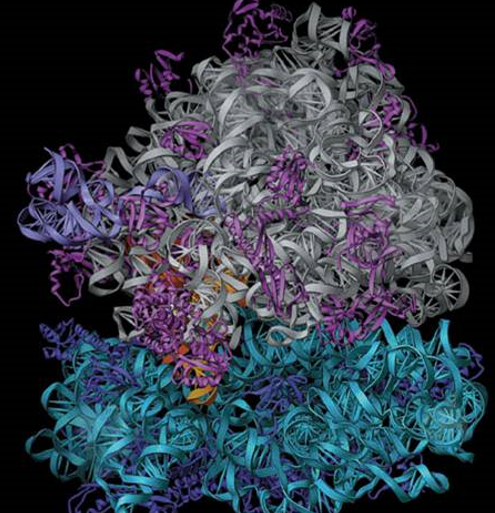
The ribosome is an organelle that is made up of two subunits with RNA and protein components. Ribosomes are also traditionally described in terms of their sedimentation coefficients.
Translation in the mRNA into a polypeptide chain is catalyzed by ribosome-specific coupling of tRNA anticodons and mRNA codons.
5.1.2.1 Binding sites in the ribosomes
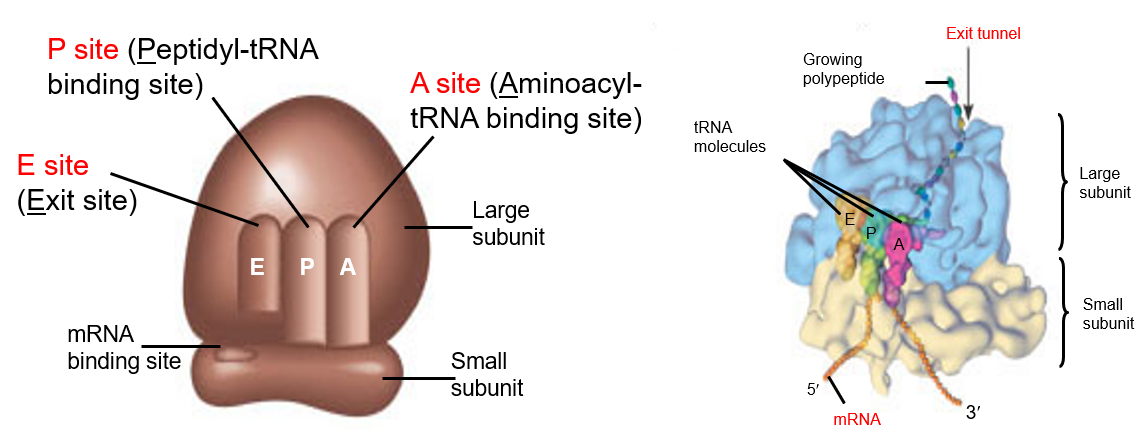
Note the three sites: the A, the P, and the E sites of the ribosomes.
5.2 Initiation, Elongation, and Termination
The below graphic summarizes the lecture’s content in a nutshell.
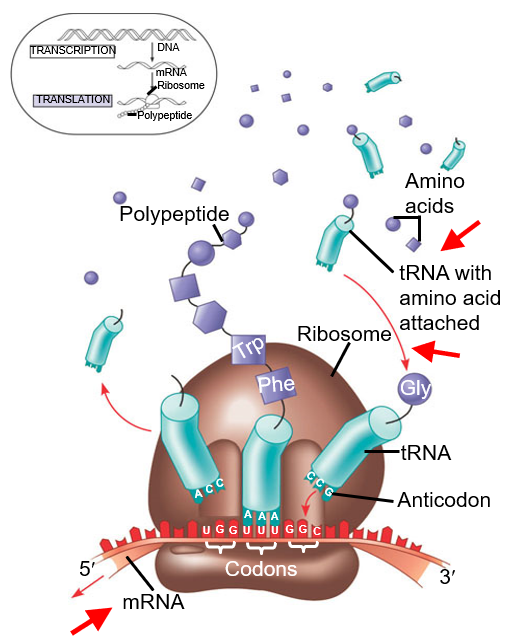
Figure 5.2: Diagram Depicting Translation
5.2.1 Protein Factors
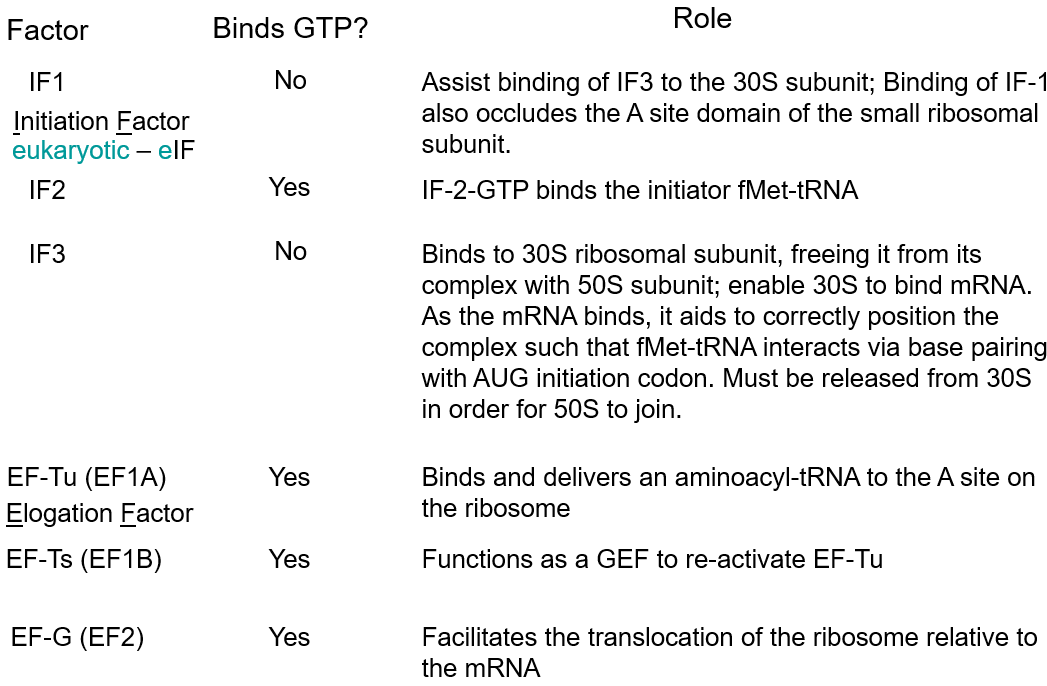
Figure 5.3: Protein Factors to Know

Figure 5.4: More Protein Factors to Know
Do memorize the content in the above slides!
5.2.1.1 GTP-binding proteins
A GTP-binding protein is a protein that has a different conformation depending on whether it is bound to a GTP or a GDP moiety. GTP stabilizes the active formation of the protein.
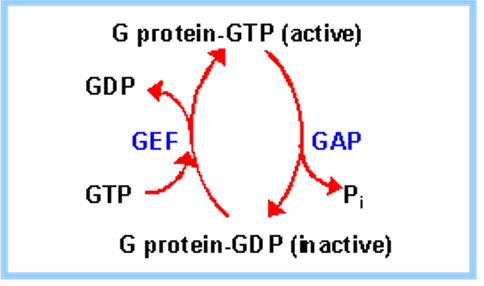
Figure 5.5: Graphic Summarizing GDP and GTP Binding in Proteins
The GTPase activating protein (i.e., GAP) hydrolyzes bound GTP to GDP + Pi.
The protein guanine exchange factor (i.e., GEF) aids in the release of GDP in exchange for GTP.
5.2.2 Initiation of translation
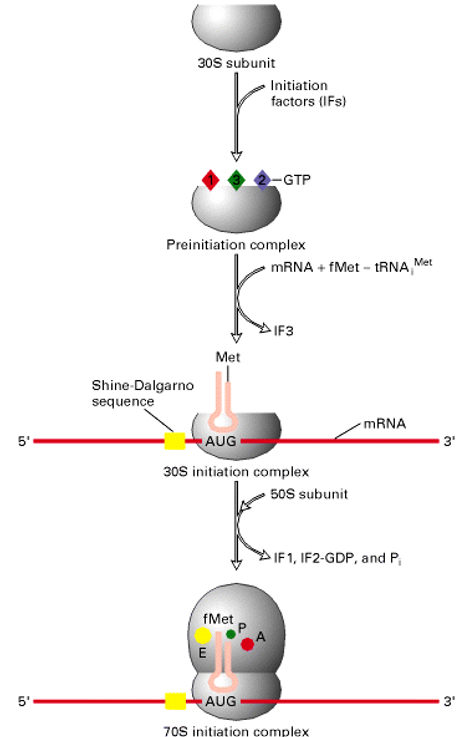
Figure 5.6: How Translation Starts
First, IF-1 binds to the 30S subunit and also assists in the binding of the IF-3. The IF-2-GTP then binds to the tRNAiMet and docks with the 30S subunit.
In the second step, the 30S pre-initiation complex binds to the mRNA and is guided by the Shine-Dalgarno sequence. It is through this sequence that the start codon AUG is located (and protein initiation will begin).
Thereafter, the 50S ribosomal subunit is added to the complex. The hydrolysis of IF-2-GTP by GAP (served by 50S) then yields the complete 70S initiation complex.
The ribosome is then ready for the entry of the next aminoacyl-tRNA at the “A” site - hence paving way for elongation.
5.2.3 Elongation
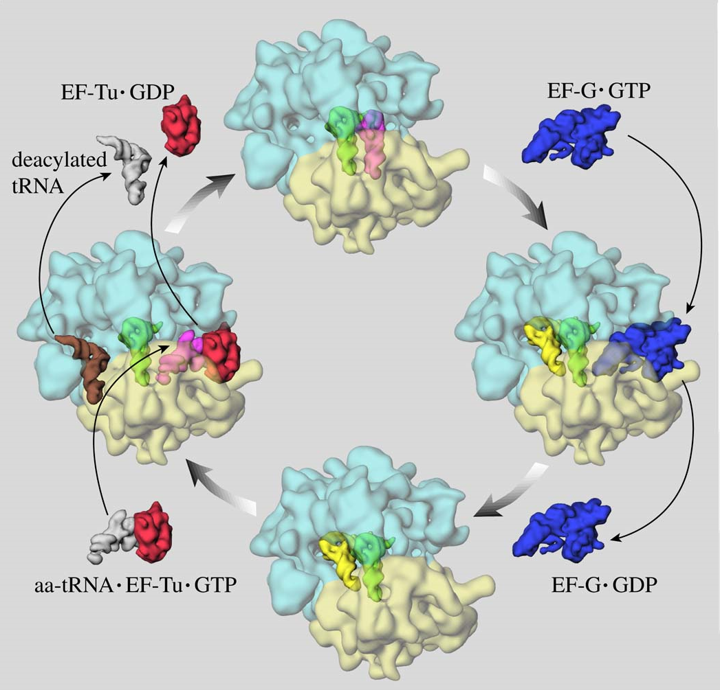
Figure 5.7: Elongation in Protein Synthesis
First, the protein factor Ef-Tu-GTP binds and delivers an aminoacyl-tRNA to the “A” site of the ribosome. The tRNA must also have the correct anti-codon to pair with the codon at the “A” site.
Note that when the tRNA binds, that it causes a conformational change to the small ribosomal subunit. This leads to conserved bases of 16S rRNA to interact (closely) with the minor groove of the first two bases of the codon / anticodon complex.
Otherwise, if the “A” site is occupied by the incorrect tRNA, then the base pair interaction will be unstable and the aminoacyl-tRNA is released (i.e., proofreading).
5.2.3.1 Role of EF-Ts in elongation
EF-Ts function as GEFs to reactivate EF-Tu - in other terms, EF-Ts help facilitate the release of GDP from EF-Tu:
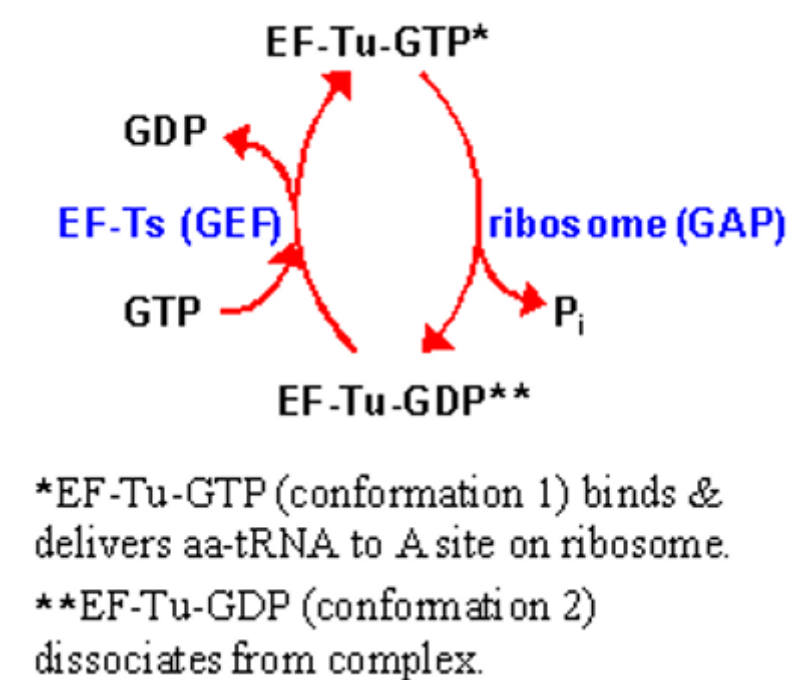
Figure 5.8: EF-Tu role in Elongation
Translocation of the ribosome (relative to the mRNA) involves the movement of EF-G-GTP (which binds to the ribosome in the vicinity of the “A” site).
Note that the binding of EF-G-GTP might push the mRNA with the attached polypeptide from the “A” site to the “P” site. In some sense, the ribosome acts as a GAP for EF-G.
5.2.4 Peptide bond formation
Transpeptidation usually involves the catalysis of acids / bases by a universally conserved adenosine of the 23S rRNA.
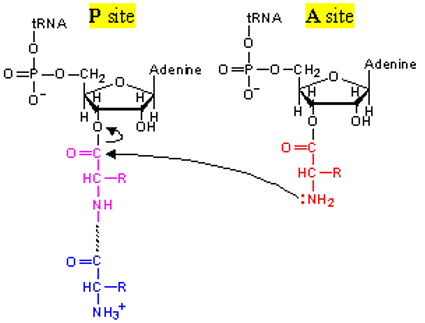
Figure 5.9: Transpeptidation
The nitrogen atom of the amino group (that is linked to the 2’ or 3’-OH of the terminal adenosine of the tRNA in the “A” site) then attacks the carbonyl carbon (see above) of the amino acid in the “P” site:
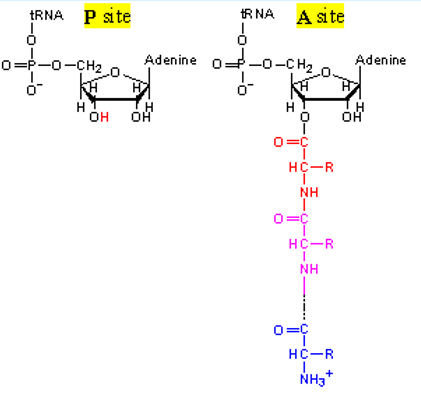
Figure 5.10: Transpeptidation (part 2)
From here on out, the translocation has already partially happened.
5.2.5 Termination
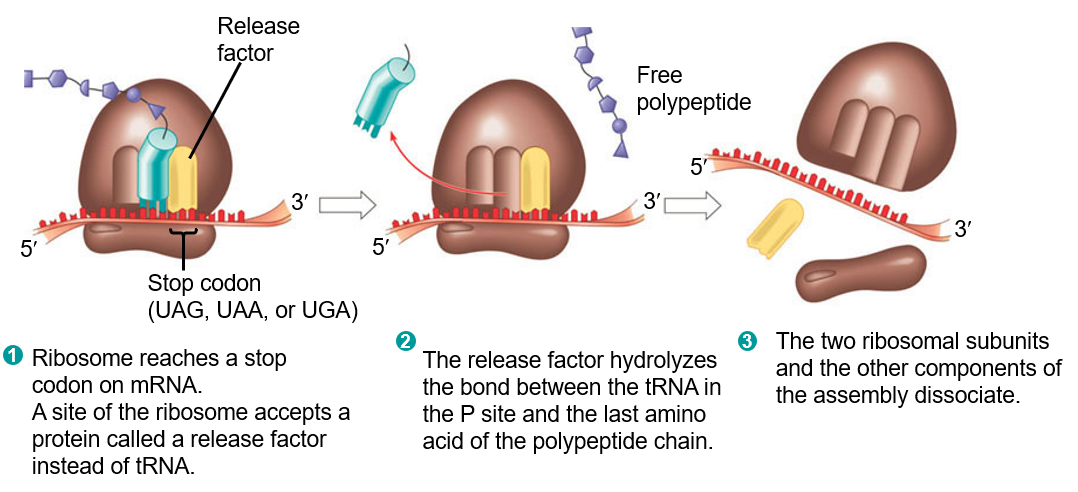
Figure 5.11: Termination of Protein Synthesis
In the first step, the ribosome must first reach a stop codon on the mRNA. When it does, a release factor is then called (instead of a tRNA).
The release factor then hydrolyzes the bond between the tRNA in the “P” site and the last amino acid of the polypeptide chain.
The above then causes the two ribosomal subunits to disassociate with one another.
5.2.5.1 Role of RFs and RRF in termination
The protein factors RF-1 and RF-2 are responsible for recognizing stop codons. RF-3-GTP facilitates the binding of RF-1 and RF-2 to the ribosome. Ribosomal peptidyl transferase then catalyzes the transfer of the peptidyl group to water
RF-3-GTP and RF-3-GDP then cause a conformational change that results in the disassociation of release factors.
A protein called the ribosomal recycling factor (i.e., RRF) is also required along with EF-G-GTP and IF-3 for the release of an uncharged tRNA from the P site and the disassociation of the ribosome from the mRNA (with the separation of the two ribosomal subunit).
5.2.6 Accuracy of translation
Protein translation is accurate for a few reasons:
There are different classes of tRNAs synthetases.
Most tRNA families are large enough to produce a wide range of specific interactions with the corresponding synthetase.
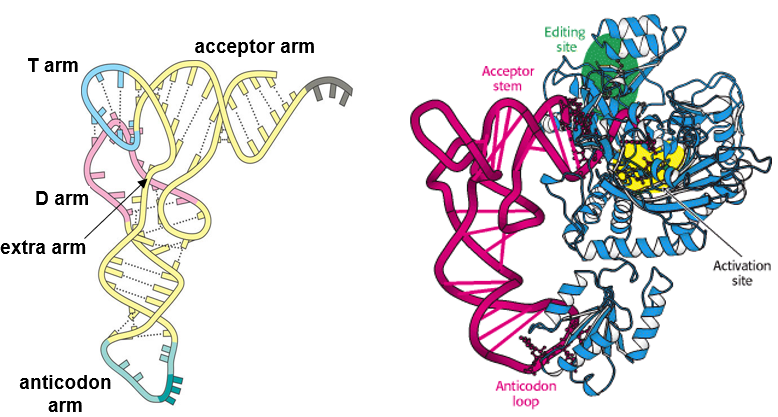
Figure 5.12: Parts of a tRNA Molecule
To be more particular, the tRNA families are large enough to produce specific interactions with the anticodon arm, the acceptor arm, and in some cases, the variable loop.
5.2.6.1 Double-sieving mechanism
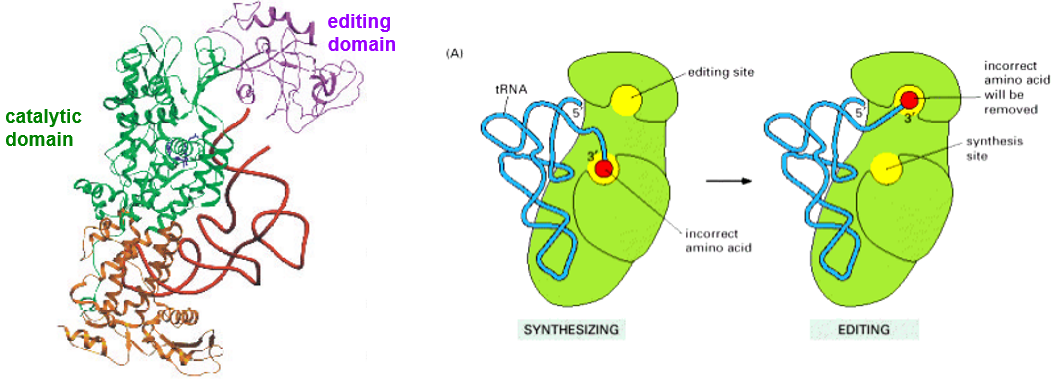
Figure 5.13: Class I Synthetases
There are two sieves to this mechanism:
In the first sieve…
The enzyme binds to a similar or a correct substrate and rejects other substrates; the correct amino acid would have a very high affinity for the active site pocket of its synthetase.
In the second sieve…
The wrong, adenylated amino acid is hydrolyzed at the 2nd editing site (see above). The wrong amino acid would be forced into the second pocket (i.e., the editing pocket) of the protein.
The tRNA molecule in both sieves is directly involved in both sieves; the CCA-end shuttles from the catalytic to the editing site via a hairpin conformation to a helical conformation.
5.2.7 Ribosomal Roles in Translational Fidelity
“We postulate that incomplete attachment will only retain the intermediate for a very brief time before the latter breaks loose and diffuses elsewhere. However, when it eventually diffuses to the correct place, it will be held by hydrogen bonds to all three bases and will thus be retained, on the average for a much longer time.”
– Crick and colleagues
Note that there are two main points highlighted in Crick et. al’s postulation:
- Time
- Base pairing
5.2.8 Specificity of base pairing
The wobble hypothesis states that the pairing between the codon and the anti-codon at the first two codon positions always follow the usual rules, but the exceptional “wobble” always shows up at the third position.

Figure 5.14: Wobble Hypothesis Illustrated
The wobbling is because of the tRNA anti-codon loop; it allows flexibility at the first base of the anticodons.
5.2.9 Induced-fit model
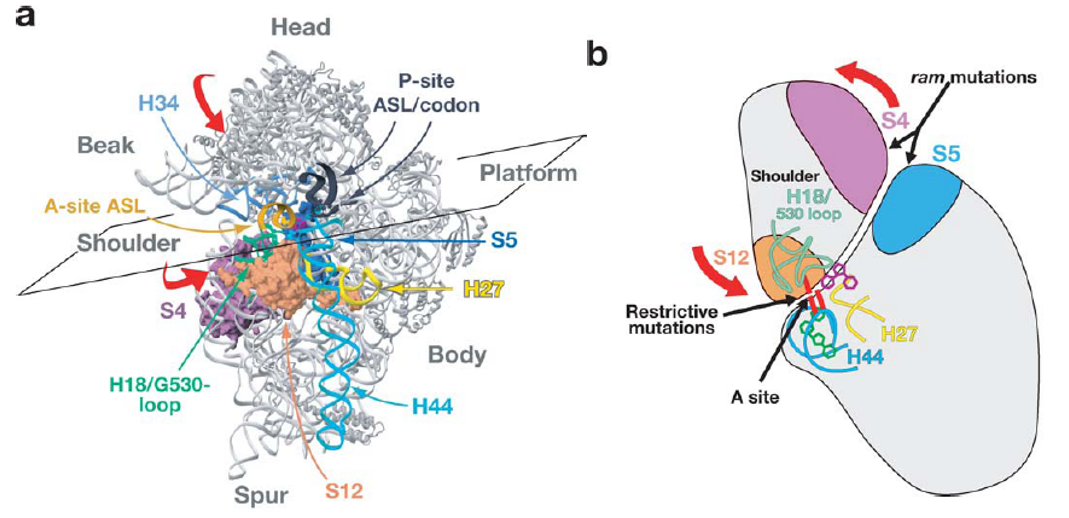
Figure 5.15: Induced Fits in the Induced-Fit Model
The model states that there is always a conformation change to the ribosome - this change is brought on by domain closure in the 30S subunit (induced by cognate tRNA binding).
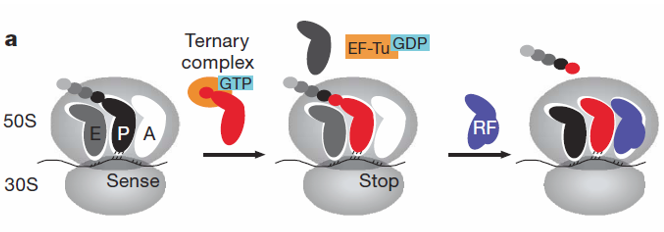
Figure 5.16: More on the Induced-Fit Model
RF-2-catalyzed reactions tend to release faster for the P-site-mismatched complex after the formation of the peptide bond (which also compromises tRNA selection fidelity).
5.2.10 Role of ribosome in translational fidelity
Here, the kinetic proofreading model is presented.
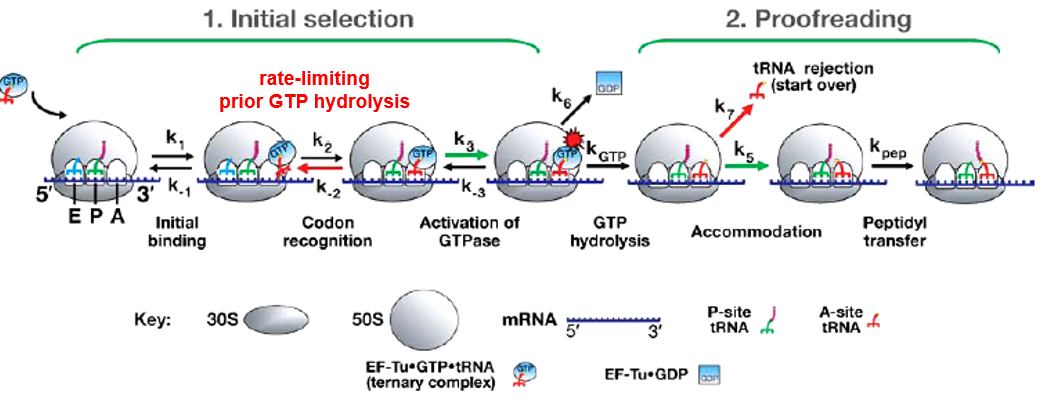
Figure 5.17: Kinetic Proofreading Model
In the codon recognition phase k2, the tRNA anticodon reversibly associates with the codon at the 30S “A” site. This leads to the stabilization of the ternary complex on the ribosome and facilitates the GTPase activation of EF-Tu (at k3, kGTP).
EF-Tu-GDP then causes a conformational change in the ribosome; either aminoacyl end of the tRNA then moves into the “A” site of the 50S ribosome (i.e., k5), which then leads to the formation of a peptide bond. Otherwise, k7 happens and the amino acid tRNA disassociates from the ribosome.
5.3 Post-Translational Modifications of Proteins
Post-translational modification refers to the chemical modification of a protein after its creation. It is also one of the later steps in protein synthesis for many proteins.
Such post-translatioanl modifications may include the following:
- Proteolytic cleavage
- Disulfide bridge formation
- Attachment of biochemical functional groups (e.g., acetate, phosphate, lipids, and carbohydrates)
However, the types of post-translational modifications we will be focusing on include the following:
5.3.1 Glycosylation
Glycsolylation is the process of adding saccharides to proteins and lipids. The majority of proteins synthesized in the rough endoplasmic reticulum undergo this process; the polysaccharides attached can serve the following functions:
- Proper folding
- Protein stability
- Protein recognition
Glycosylation is an enzyme-directed site-specific process as opposed to the non-enzymatic chemical reaction of glycation; it can also be N-linked or O-linked.
5.3.1.1 Dolichol
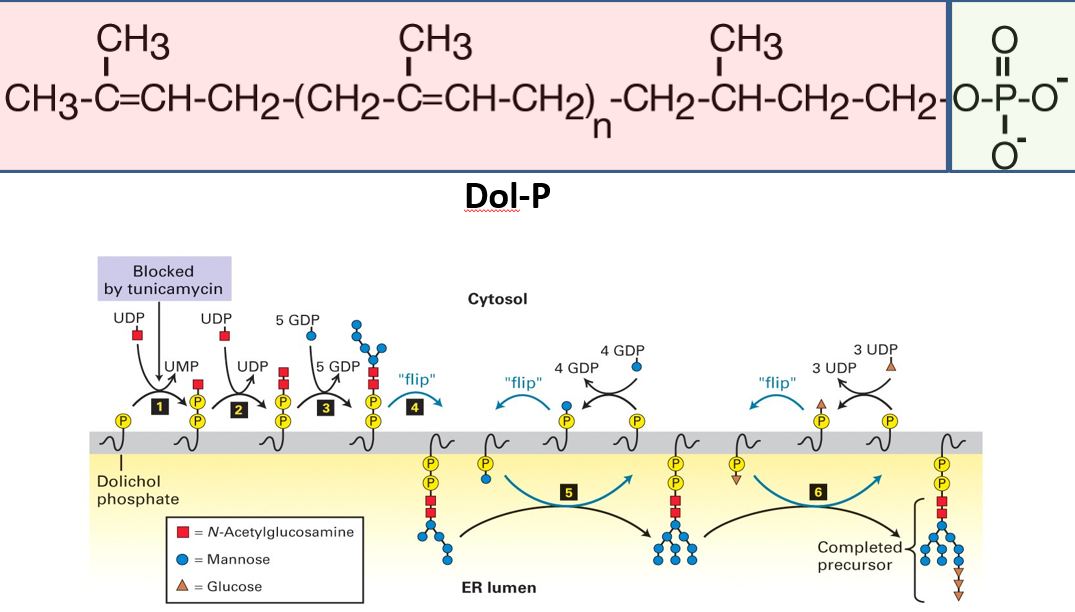
Figure 5.18: Structure and Location of Dolichol
Dolichol is a polyisoprenol lipid; it is made up of five-carbon isoprene units that have been linearly linked in a head-to-tail fashion (see above).
This lipid is embedded in the lipid bilayer.
5.3.1.2 N-linked glycoproteins
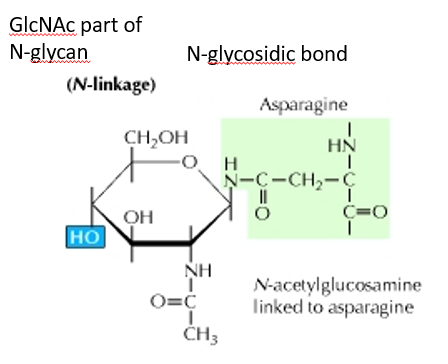
Figure 5.19: A N-Linked Glycoprotein
Here, the carbohydrate group is attached to the N atom of the side chain of asparagine.
The sugar that is attached is typically N-acetylglucosamine (i.e., GlcNAc)
5.3.1.3 O-linked glycoproteins
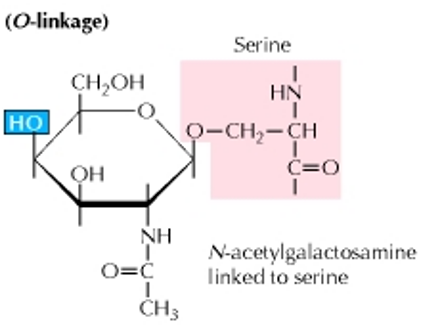
Figure 5.20: An O-Linked Glycoprotein
In contrast to an N-linked glycoprotein, the carbohydrate is attached to the O atom of a serine or a threonine molecule.
The sugar that is attached to the O atom is typically N-acetylgalactosamine.
5.3.2 Phosphorylation
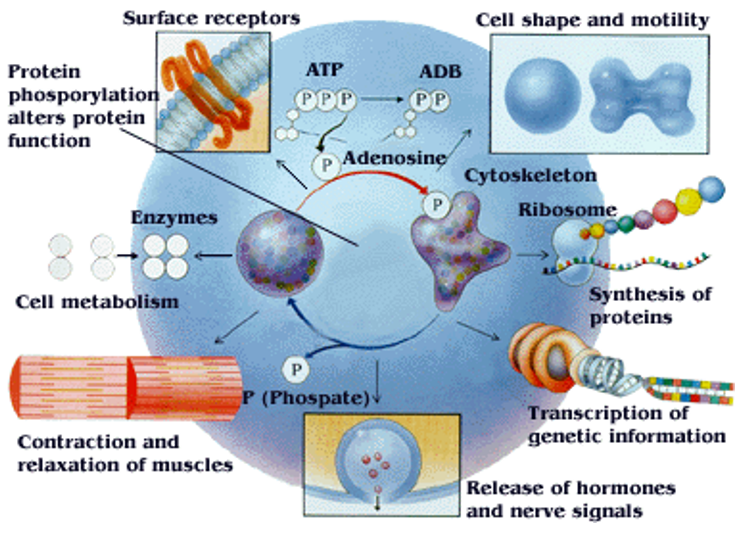
Figure 5.21: Phosphorylation as a Regulatory Process
In 1992, scientists Edmond H. Fischer and Edwin G. Krebs discovered reversible protein phosphorylation as a means of regulating biological mechanisms.
Nonetheless, phosphorylation is the addition of a phosphate (i.e., PO4) to a protein’s serine, tyrosine, or a threonine moiety.
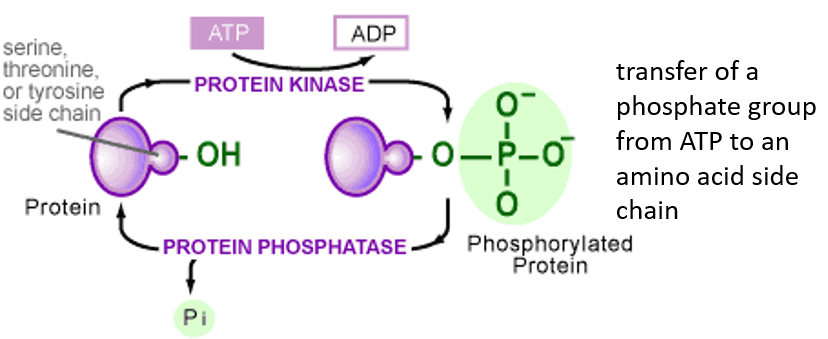
Figure 5.22: Activity of a Kinase and a Phosphatase
A phosphatase removes phosphate groups; a kinase does the exact opposite.
5.3.2.1 Phosphorylation in signal transduction
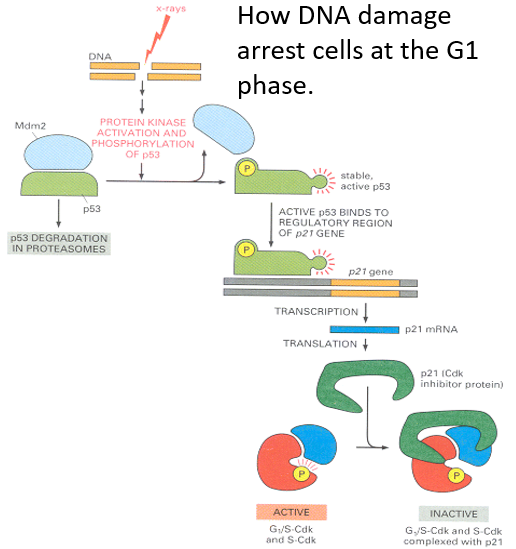
Figure 5.23: Phosphorylation During Mitosis in the S Phase
Rb phosphorylation is a mechanism that controls the initiation of the S phase in animal cells.
When lamins are phosphorylated during mitosis, this results in the breakdown and subsequently, the re-formation of the nuclear envelope during mitosis.
5.3.3 Acetylation
Acetylation involves adding an acetyle group to an organic compound (typically happens at a Lysine residue).
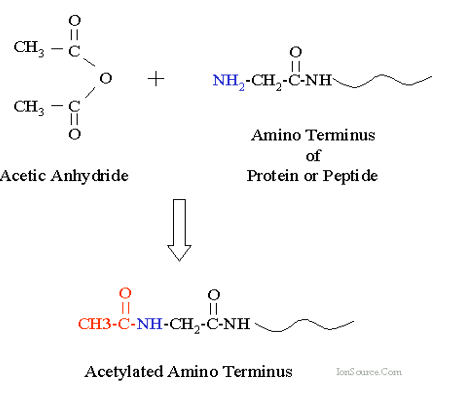
Figure 5.24: Acetylation at Lysine
For instance, histones are first acetylated and deacetylated on their lysine residues as part of gene regulation. These reactions are typically catalyzed by enzymes called histone acetyltransferase (i.e. HAT) or histone deacetylase (i.e., HDAc). The molecule acetyl CoA provides the source of acetyl groups during acetylation.
5.3.3.1 Histone (de)acetylation
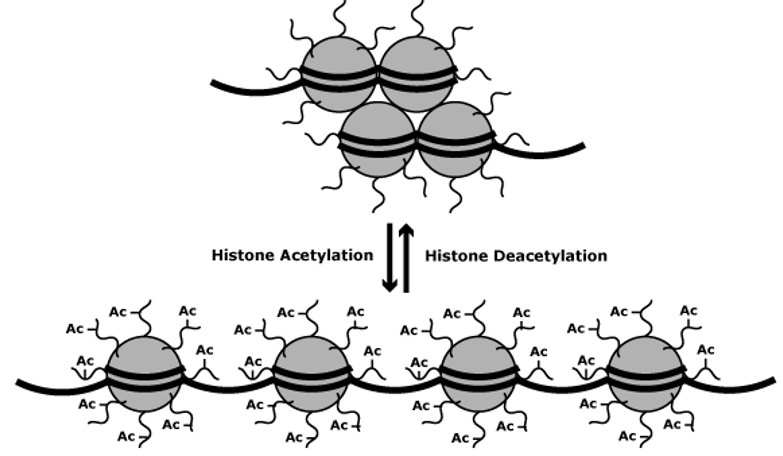
Figure 5.25: Impact of Acetylation and Deacetylation on Gene Expression
Acetylation of lysine residues at the N-terminus of a histone protein removes positive charges (hence also reducing the affinity between DNA and histones). Because of this, RNA polymerase and transcription factors can easily access the promoter region.
In most cases, acetylation enhances transcription whereas deacetylation represses transcription.
5.3.4 Methylation
Methylation is the replacement of a hydrogen atom with a methyl group. This typically occurs on arginine or lysine residues in the protein sequence.
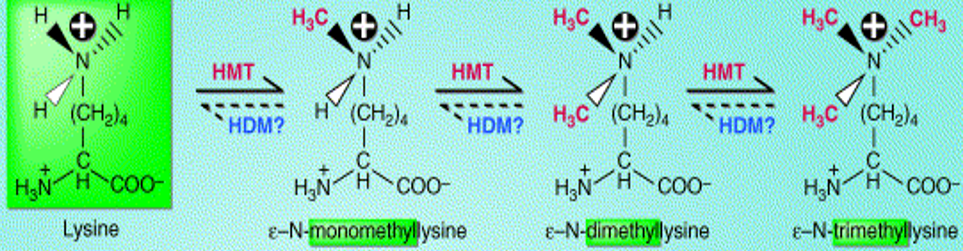
Figure 5.26: Methylation of a Lysine Molecule
Methylation is best studied in histones. Transferring methyl groups from S-adenosyl methionine is an action catalyzed by enzymes known as histone methyltransferases (i.e., HMTs).
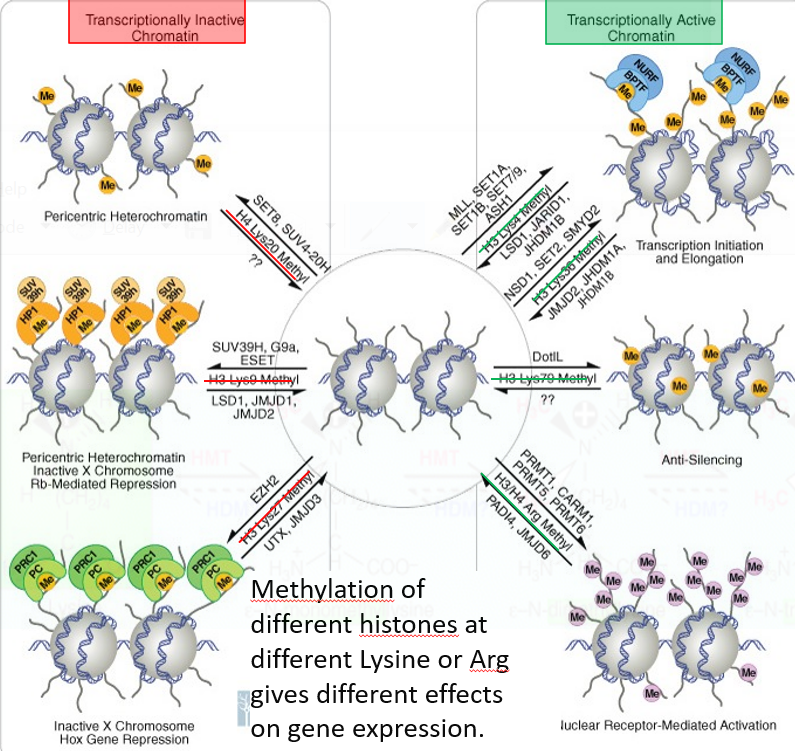
Figure 5.27: Effect of Methylation on Chromatin
Nonetheless, histones that are methylated on certain residues can also serve to activate or repress gene expressions.
5.3.5 Signal transduction
Nuclear hormone receptors (i.e., NRs) are part of a superfamily of ligand-activated transcription factors that regulate the expression of genes that play roles in processes like metabolism, growth, and reproduction.
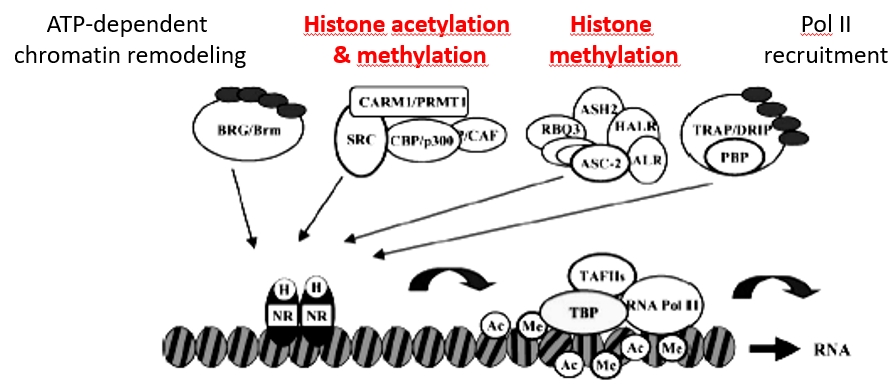
Figure 5.28: Acetylation and Methylation in Signal Transduction
NRs also mediate transcriptional response to hormones like sex steroids, the adrenal steroid, vitamin D3, thyroids, and retinoids hormones in addition to a variety of metabolic and dietary lipids.
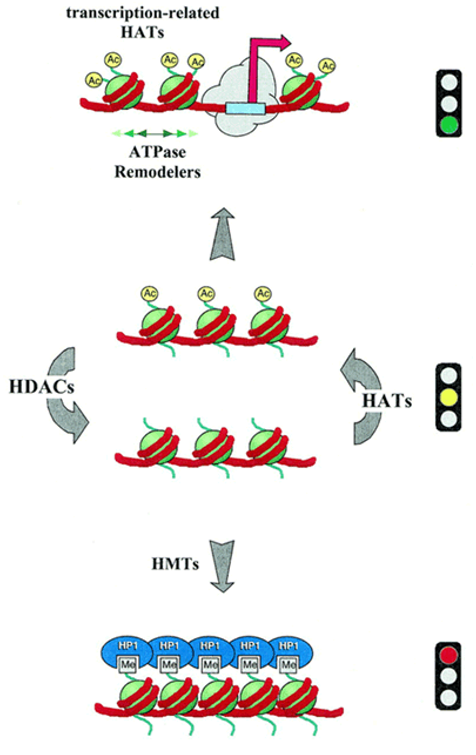
Figure 5.29: Activities of HAT and HDAc in the Status of Chromatin
HAT1 and HDAc activities can also help negotiate the acetylation status of chromatin (recall that acetylation permits ATP-dependent chromatin factors that are open to promoters).
Deacetylation (which is followed by histone methylation) may also form a base for highly repressive structures such as heterochromatin.
5.3.6 Ubiquitination
In 2004, scientists Aaron Ciechanover, Avram Hershko and Irwin Rose were given the nobel prize in chemistry for their discovery of the ubiquitin system.
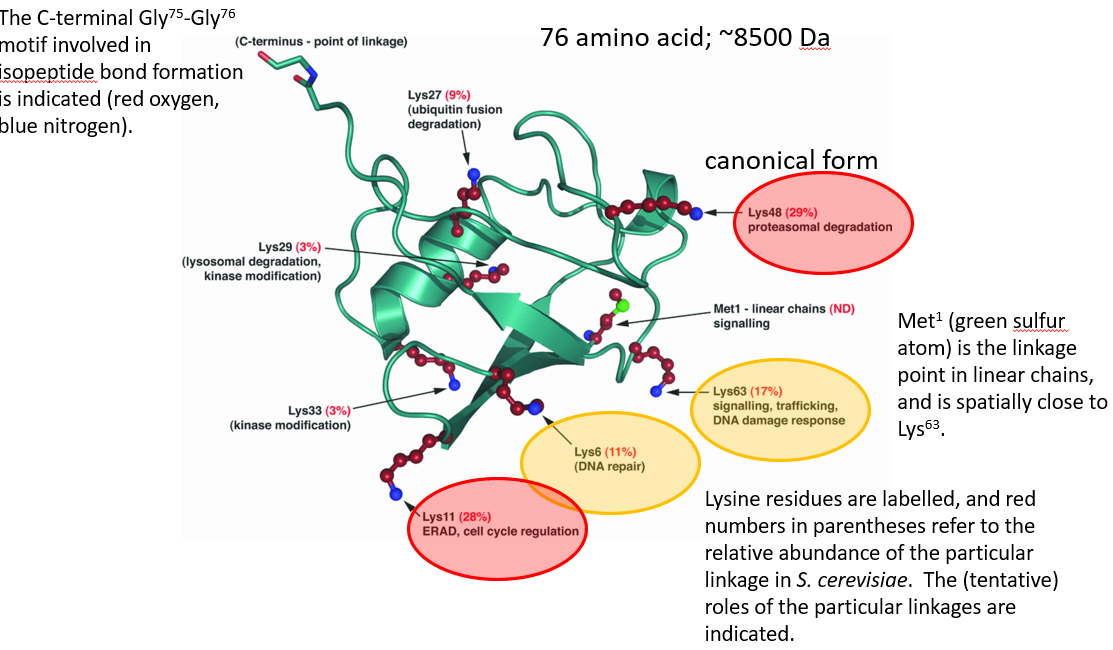
Figure 5.30: Structure of Ubiquitin
Ubiquitination is a cellular regulatory process where ubiquitins - a kind of protein - are added to a larger protein: the target protein.
Ubiquitin is generally attached via C-terminal glycine residue to the \(\epsilon\)-amino group of a lysine residue. The reaction is performed by a sophisticated three-step enzymatic cascade.
In many human diseases (e.g., cancer and infections by pathogens), the de-regulation of ubiquitin-mediated signalling is implicated.
5.3.6.1 Forms of ubiquitination
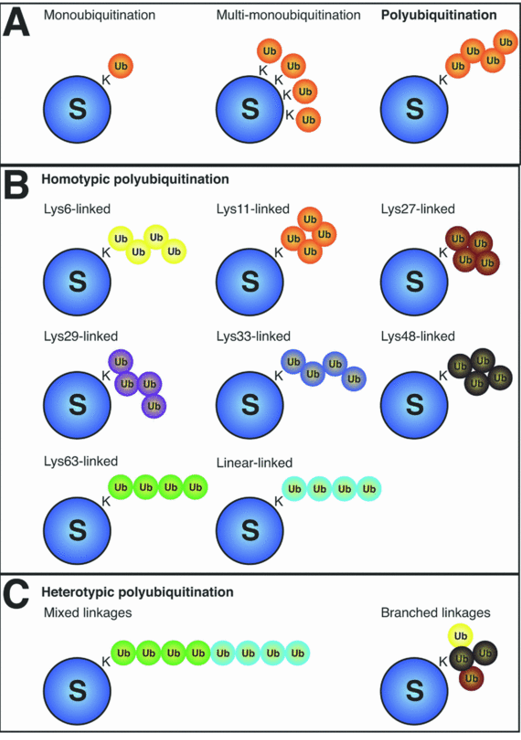
Figure 5.31: Types of Ubiquitin
Homotypic polyubiquitination occurs when each ubiquitin chain contains a single linkage type.
Heterotrophic polyubiquitination happens when there are mixed, alternating linkage types. In branched or forked polyubiquitin chains, a single ubiquitin protein is extended at two or more amino acid residues.
5.3.6.2 How does ubiquitination happen?
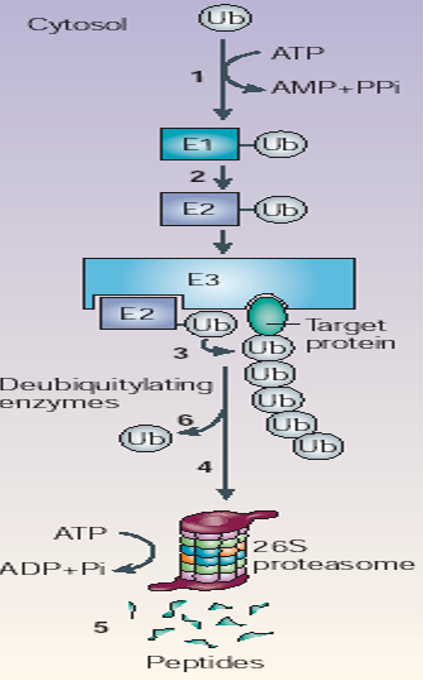
Figure 5.32: Steps of Ubiquitination
In the first step (ATP-dependent), an E1 ubiquitin-activation enzyme charges an E2 ubiquitin conjugating enzyme with ubiquitin; it links ubiquitin to E2’s catalytic cysteine residue via a thioester linkage.
In E3, ubiquitin ligases function as adaptors that bind both substrate and charged E2 proteins. E3 ligases then facilitate isopeptide bond formation between ubiquitin and the substrate by complicated mechanisms!
Polyubiquitination occurs because ubiquitin molecules are attached to lysine residues in ubiquitins that have already been attached. Also note that polyubiquitination substrates are bound to the proteasome.
5.3.7 Dealing with abnormal proteins
The proteasome is the cell’s so-called waste disposer. It is also the central enzyme of “non-lysomal protein degradation”. The 26S proteasome 19S lid and base and a 20S proteasome component.
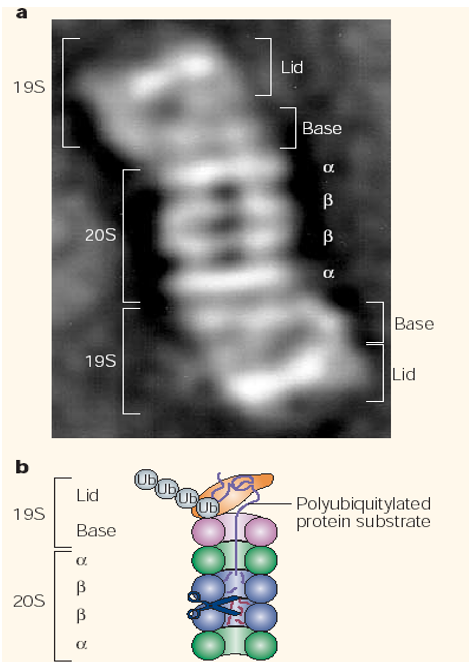
Figure 5.33: Structure of a Proteasome
The 20S component is a cylinder with the active sites in its interior. The 19S particle has a base of six ATPase rings that is associated with the 20S particle.
5.3.7.1 Ways of dealing with abnormal proteins and abnormal proteins.
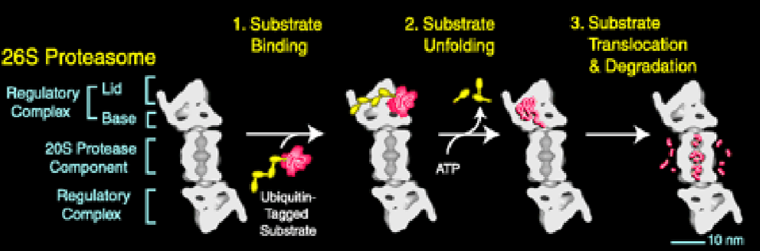
Figure 5.34: How the Proteasome Works
The polyubiquitin chain is removed by isopeptidases from the substrate before its movement into the proteolytic chamber.
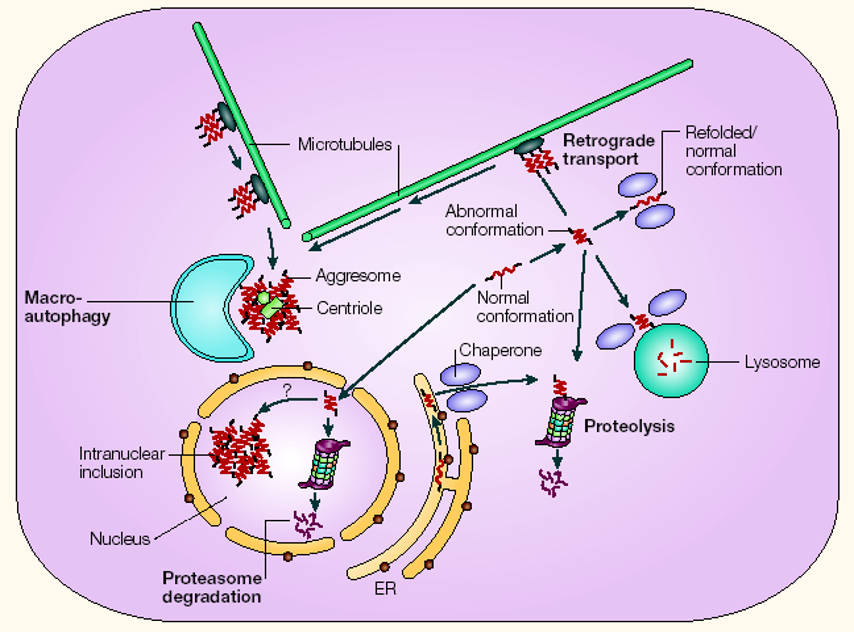
Figure 5.35: Ways a Cell Deals with Abnormal Proteins
There are many ways that a cell can deal with abnormal proteins.
HAT = Histone Acetyltransferase↩︎