Topic 2 Mitochondrial and Plastid Genomes
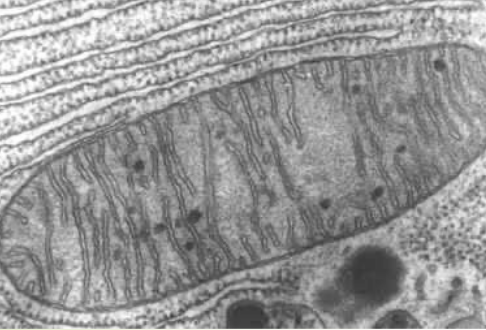
Figure 2.1: Electron Microscopy of a Mitochondrion
Mitochondria are organelles that serves as cells’ energy sources. They convert (potential) energy from food into adenosine triphosphate (i.e., ATP). Mitochondria also have an inner membrane (i.e., the cristae) and an outer membrane (i.e., the part that limits the organelles), constantly change their shapes due to fission and fusion, and are often shown as rod-shaped figures:
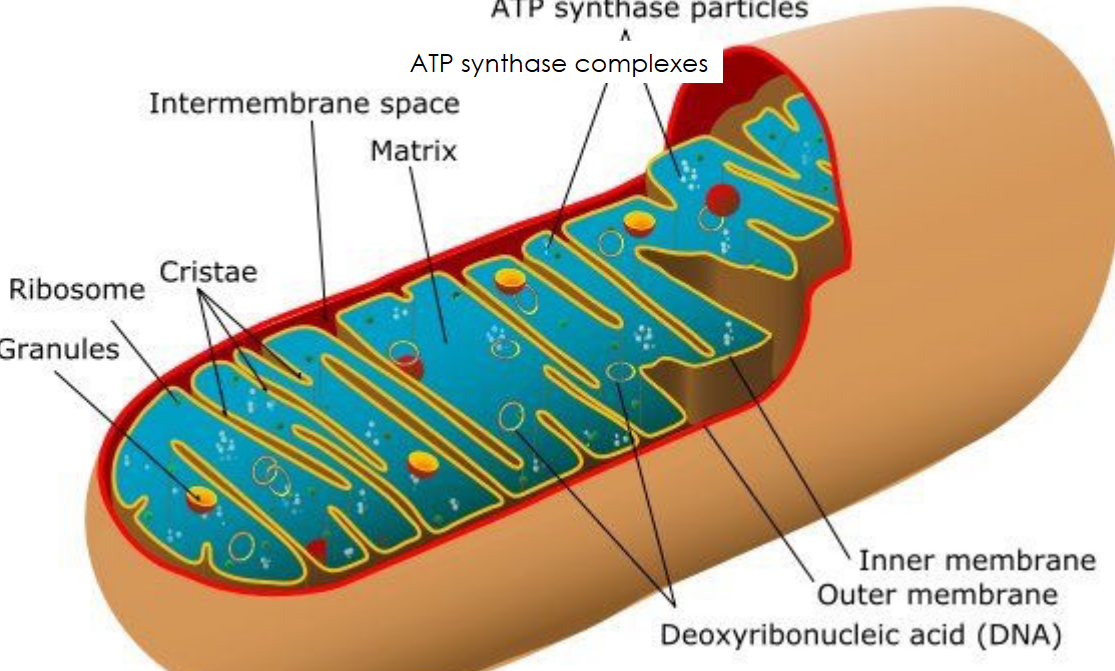
Figure 2.2: Anatomy of a Mitochondria
For obvious reasons, cells that require tons of energy (e.g., muscle cells) also have many mitochondria. It is also interesting to note that the Trypanosome protozoan has one large mitochondria.
Human liver cells have 1000 to 2000 mitochondria each; the organelle can also occupy up to 25% of the cell’s cytoplasm.
2.1 Parts of the Mitochondria
2.1.1 Outer membrane
This part of the mitochondria contains porins: channels that allow molecules and ions to move in and out of the mitochondrion.
The outer membrane is permeable to molecules that are 5000 daltons or less in size (including small proteins). Because of this, the inter-membrane space and the cytosol also contain similar concentrations of small solutes and proteins.
2.1.2 Inner membrane
This part of the mitochondrion is made up of five integral (membrane) proteins that form the respiratory chain complexes:
- NADH Dehydrogenase (i.e., Complex I)
- Succinate Dehydrogenase (i.e., Complex II)
- Cytochrome C Reductase (i.e., Complex III)
- Cytochrome C Oxidase (i.e., Complex IV)
- ATP Synthase (i.e., Complex V)
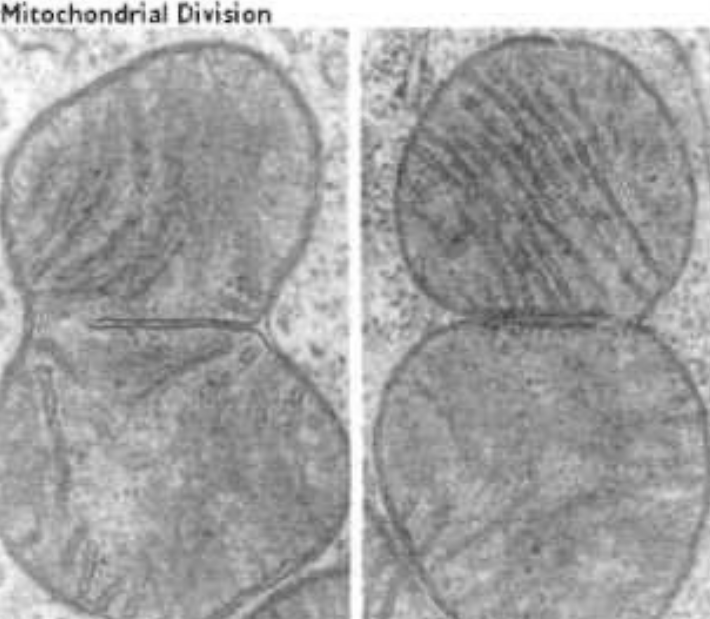
Figure 2.3: Mitochondrion Dividing via Fission
Note that other proteins in the inner membrane include (but are not limited to) translocases and proteins that are involved in mitochondrial fission and fusion.
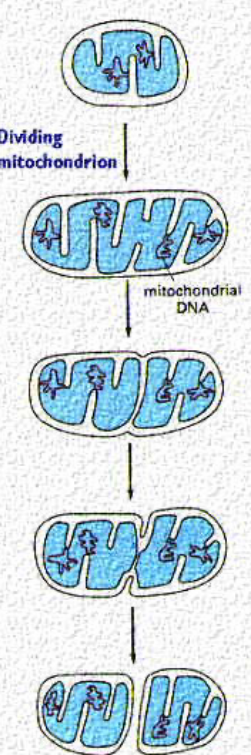
Figure 2.4: Fission and Fusion in a Mitochondrion
While all mitochondria replicate like bacterial cells, mitochondria will only divide via fission once they have reached a certain size (see the mitochondria in the middle).
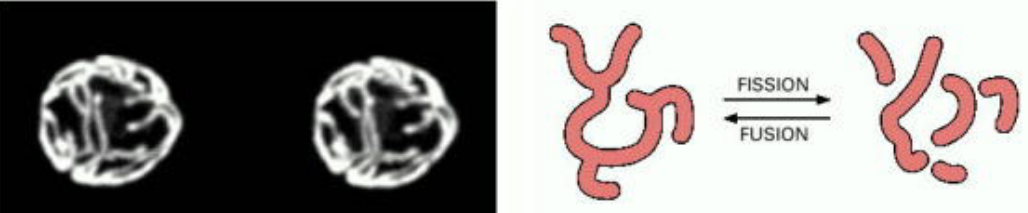
Figure 2.5: A Close-Up of Fission and Fusion in a Mitochondrion
This balance of fission and fusion also determines the arrangement of mitochondria in different cells.
2.1.3 ATP production
Glycolysis is the decomposition of glucose into two molecules of pyruvate. This is a process that occurs outside of the mitochondria.
The Kreb’s cycle then decomposes acetyl coenzyme A (i.e., acetyl-CoA) into carbon dioxide and NADH - this cycle occurs on the surface of the mitochondrion.
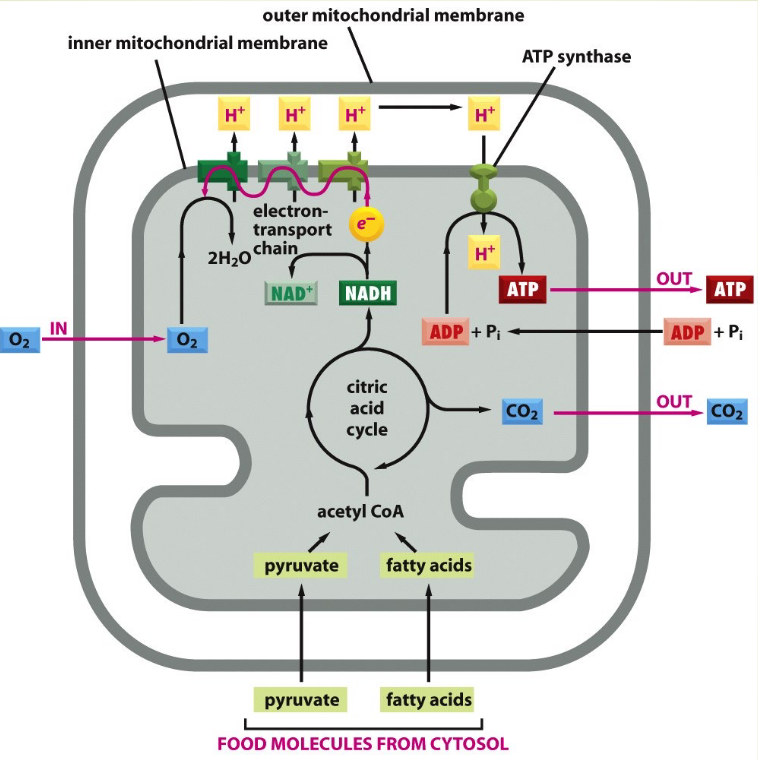
Figure 2.6: Schematic of ATP Generation
While some ATP is generated during the Krebs cycle, the large majority of ATP generation occurs during the electron transport chain. The inner surface of the cristae contains enzymes that are capable of synthesizing ATP from ADP and phosphate.
2.2 Plastids
Plastids are structures that form from proplastids: small organelles found in plant cells.
Several kinds of plastids (e.g., chloroplasts, chromoplasts, and amyloplasts) develop from proplastids and have a different function each. Undifferentiated proplastid cells are found in the meristemic cells of the plant’s shoot, and the plant’s root can differentiate into a number of highly specialized plastid types.
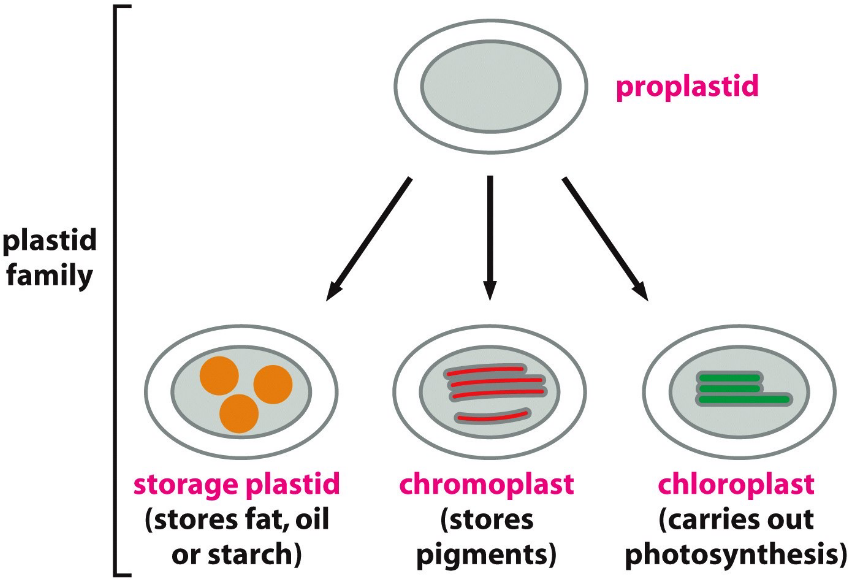
Figure 2.7: Different Types of Plastids
Green chloroplasts are found in mature mesophyll cells, colorless amyloplasts in roots and tubers, and colored chromoplasts in fruits.
Chloroplasts are the most common plastids. Each plant cell has up to several hundred chloroplasts.
2.2.1 Chloroplasts
These harvest energy from sunlight and use the energy to convert CO2 and H2O to sugars. This process is known as photosynthesis.
A typical plant cell has close to 50 chloroplasts.
2.2.1.1 Structure of a chloroplast
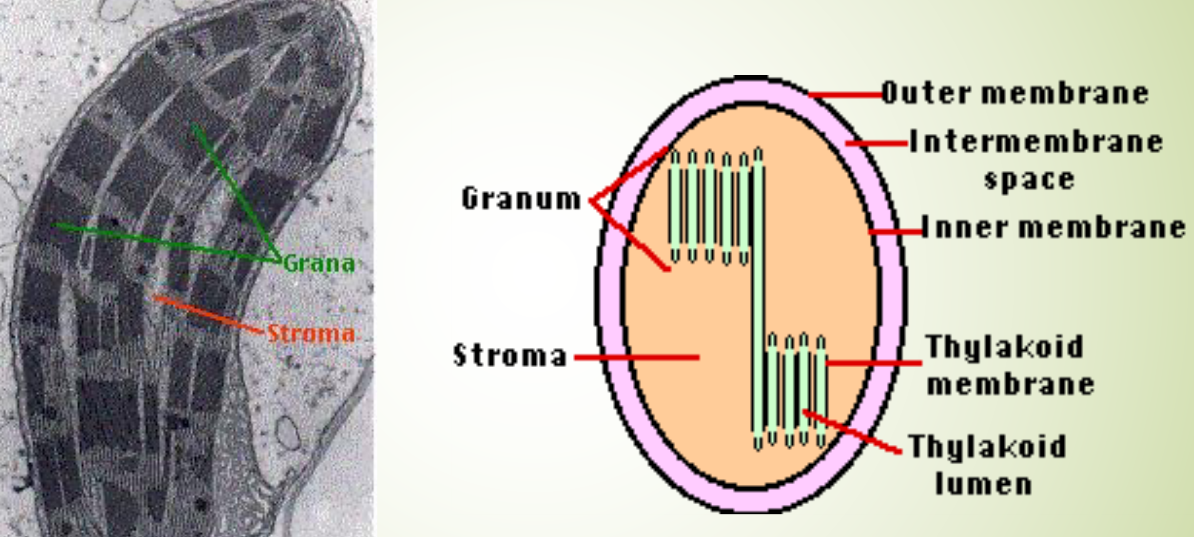
Figure 2.8: Inner Structures of a Mitochondrion
There are three different membranes to take note of:
- A smooth outer membrane that is freely available to molecules.
- A smooth inner membrane that contains transporters: integral membrane proteins that regulate the flow of materials into and out of the chloroplast.
- A system of thylakoid membranes.
2.2.1.2 Thylakoid membranes
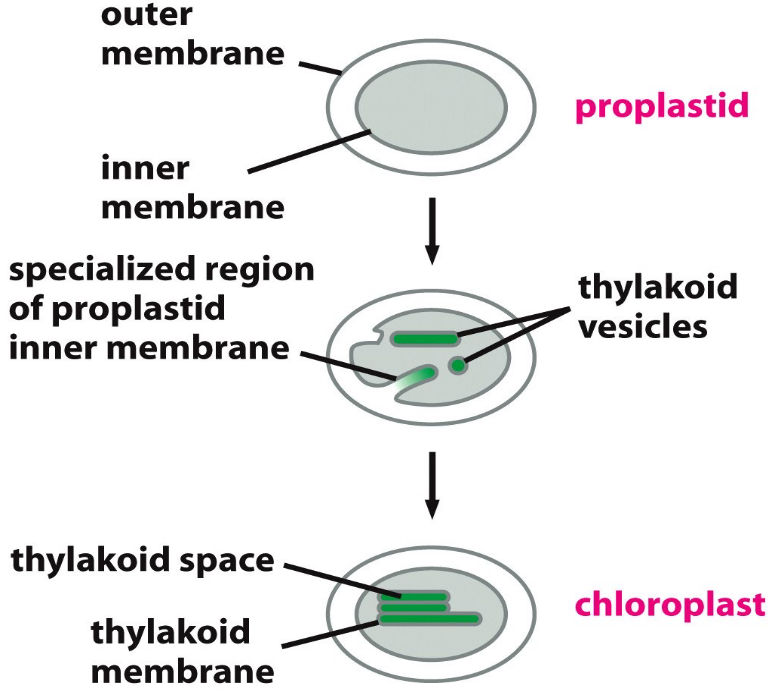
Figure 2.9: Development of the Thylakoid Membranes
The thylakoid membranes enclose a lumen: a system of vesicles (that may all be interconnected). The membranes are also surrounded by a fluid stroma. The stroma contains:
- Enzymes that participate in the dark reactions of photosynthesis.
- A special set of ribosomes, RNAs, and chloroplast DNAs.
Grana are stacks of thylakoid disks - these are located in certain places of the chloroplast.
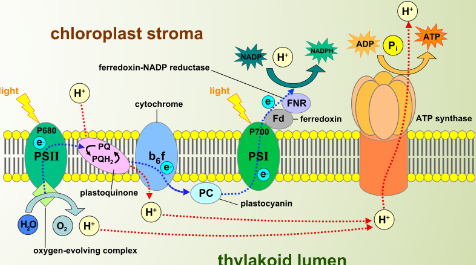
Figure 2.10: Protein Assemblies on the Thylakoid Membranes
There are four kinds of protein assemblies that are embedded in the thylakoid membranes:
- Photosystem I
- Photosystem II
- Cytochrome b6f complex
- ATP synthase
The thylakoid membranes are responsible for the light-dependent reactions of photosynthesis.
2.2.1.3 Tranport complexes on the outer and the inner membranes
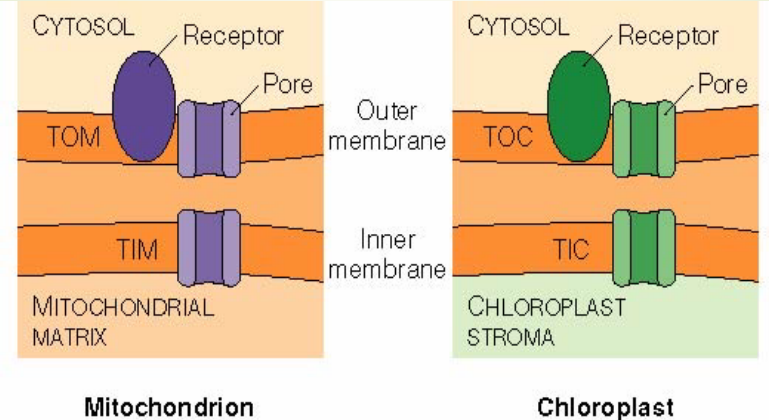
Figure 2.11: Protein Complexes on the Outer and the Inner Membranes
Note that:
- TOM: Translocase on the *outer m**itochondrial membrane
- TIM: Translocase on the inner mitochondrial membrane
- TOC: Translocase on the outer chloroplast membrane
- TIC: Translocase on the inner chloroplast membrane
2.2.1.3.1 An experimental setup to observe mitochondrial imports
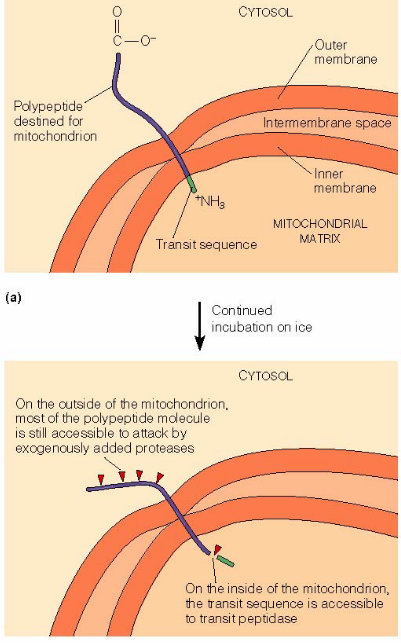
Figure 2.12: An Experimental Setup to Observe Susbtances Imported into the Mitochondrial Membranes
First, the cell-free import system was incubated on ice.
At lower temperatures, polypeptide translocation stops. The transit sequence at the N-terminus is cleaved by the transit peptidase in the mitochondrial matrix.
Proteolytic enzymes that are added to the outside of the mitochondrion will leave the C-terminus part of the polypeptide chain.
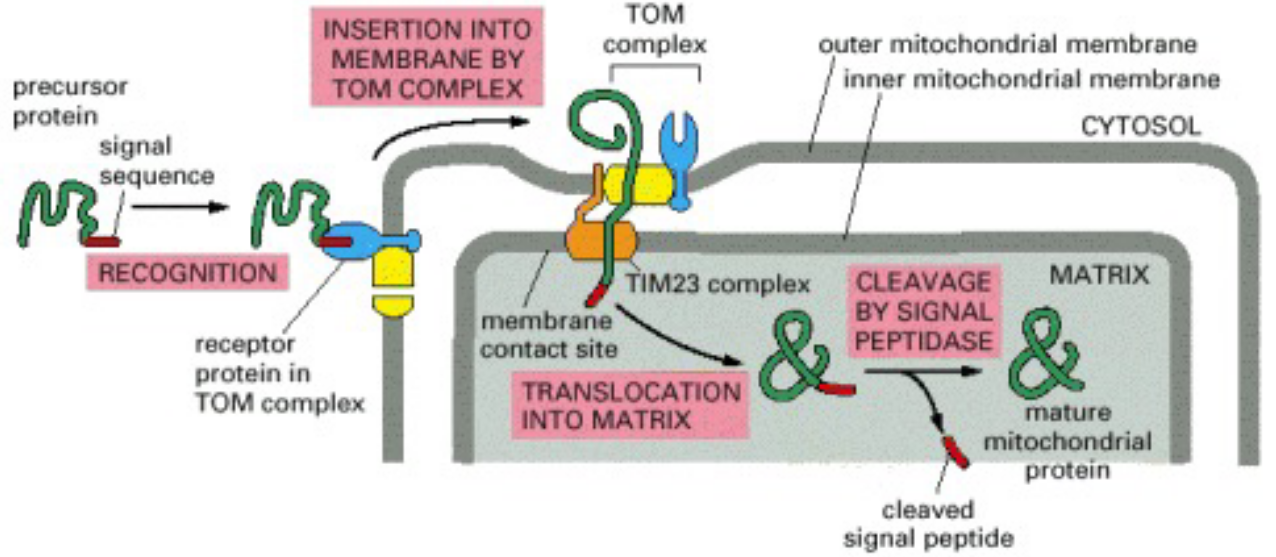
Figure 2.13: Schematic of Protein Import into the Mitochondria
The above graphic also provides a visual reference of the material discussed in the current and the previous sub-topics.
2.2.2 Comparisons between a mitochondrion and a chloroplast
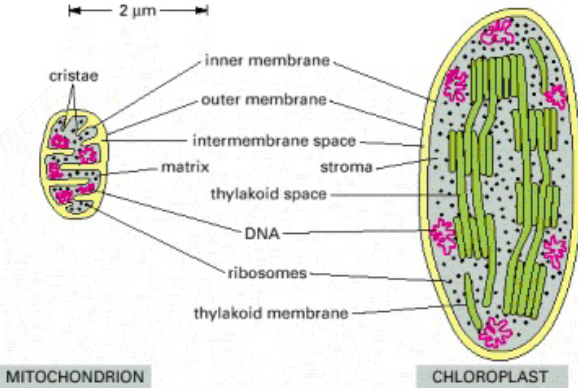
Figure 2.14: Side-by-Side Comparison of a Chloroplast and a Mitochondrion
Chloroplasts are generally larger. Furthermore, chloroplasts have a thylakoid membrane.
The inner membranes of the mitochondria are folded into cristae, and a mitochondrion imports most of its lipids. The chloroplast makes most of its own.
The mitochondrial genome is about 6000 to 300000 base pairs big. The chloroplast genome is about 70000 to 200000 base pairs big.
2.3 DNA Replication in Mitochondria and Chloroplasts
In most cells, the DNA replication of an organelle happens in all stages of the cell cycle - hence the term out-of-phase replication.
Fortunately, this process is regulated under constant conditions to ensure that the total amount of organelle DNA molecules doubles in every cell cycle.
2.3.1 Mitochondrial DNA
Because mitochondria have their own DNA and their own ribosomes, they are able to make some of their proteins. However, they still require nuclear DNA for other proteins.
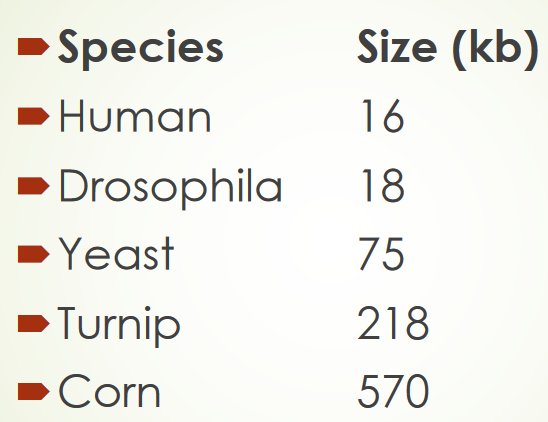
Figure 2.15: Varying Mitochondrial DNA Sizes
Most mitochondrial genomes are circular and there is a large size discrepancy among different species (e.g., plants and animals). Human mitochondrial DNA encodes for 13 proteins - the 22-fold larger Arbidopsis mitochondrial DNA encodes for 32 proteins.
Hence, the extra DNA carried by larger genomes do not appear to be necessary for function. If anything, studies have revealed that much of this DNA is made up of repeat sequences.
2.3.1.1 Human mitochondrial genome
This contains five circular mitochondrial DNA, each one about 16595 base pairs big. The DNA encodes for:
- 2 ribosomal RNA molecules
- 22 transfer RNA molecules
- 13 polypeptides
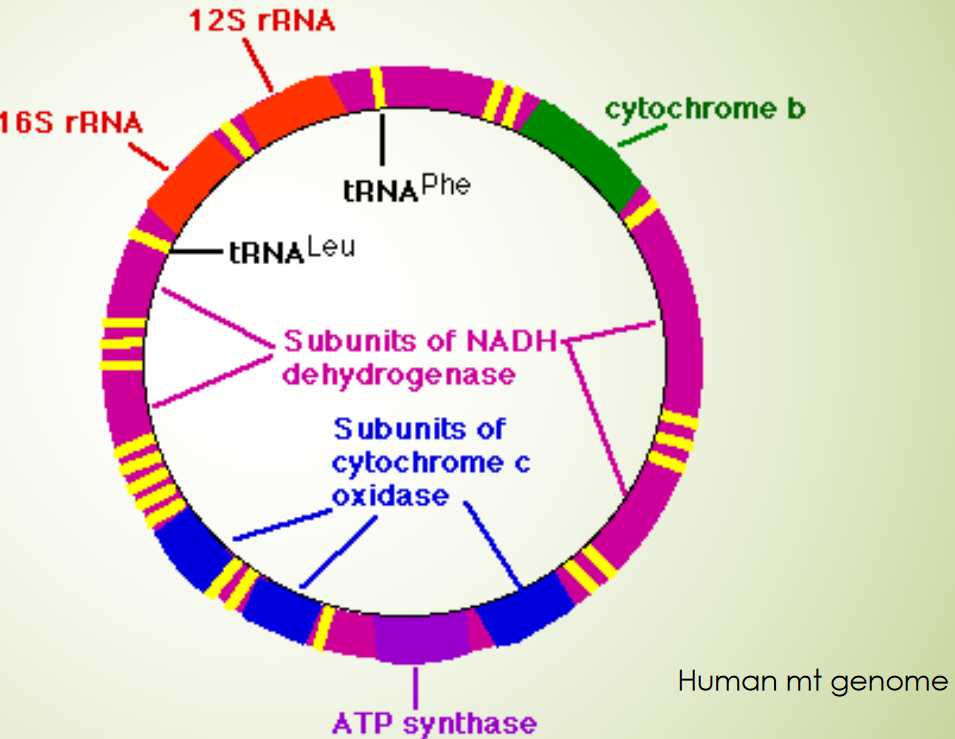
Figure 2.16: Human Mitochondrial DNA Map
The 13 polypeptides also partake in building several protein complexes:
- 7 subunits for NADH dehydrogenase
- 3 subunits for cytochrome c oxidase
- 2 subunits for ATP synthase
- 1 subunit for cytochrome b
Human mitochondrial DNA appears to be densely packed. Nearly every nucleotide appears to be part of a coding sequence (either for protein, tRNAs, or rRNAs). There doesn’t appear to be any regulatory sequences.
There are two additional features to beware:
Relaxed codon usage
30 or mote tRNAs specify amino acids in the cytosol or the chloroplasts, but only 22 tRNAs are required for the mitochondria.
Variant genetic code
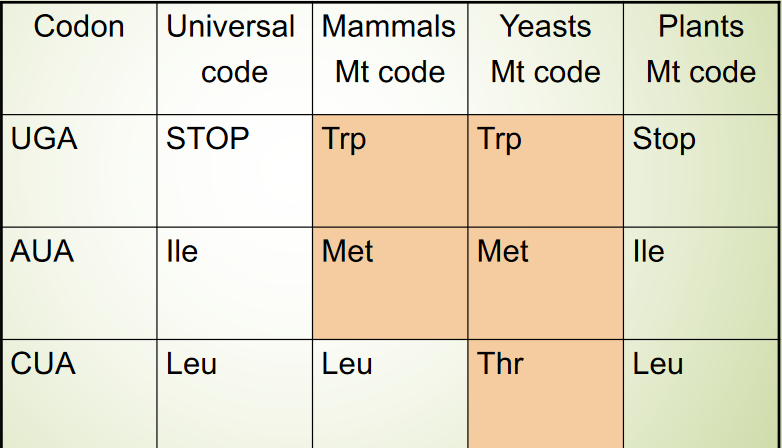
Figure 2.17: Variant Mitochondrial Genetic Codes
Some codons have different “meanings” from those of the same codons in other genomes. The codons AGA and AGG code are stop codons!
2.3.1.2 Mitochondrial inheritance
In mammals, mitochondria are inherited from the mother. Sperms carry about 100 mitochondria (most of which are located at the midpiece of the sperm and might be lost during fertilization) while oocytes have about 100000 mitochondria. As cells divide, a larger amount of the paternal mitochondrial DNA are diluted out.
A recent study conducted in the model organism C. elegans realized that mitochondrial endonuclease G decomposes paternal mitochondrial DNA upon fertilization and leads to autophagy.
2.3.1.3 Biparental inheritance
17 individuals in three separate multi-generational family exhibit biparental inheritance.
2.3.1.4 Mitochondrial genome mutations
Mutations in the mitochondrial DNA can cause diseases of all sorts, including but not limited to:
- Leber’s hereditary optic neuropathy
- “Ragged muscle fibers” associated with jerky movements. This is associated with a mutation of mitochondrial lysine tRNA.
- Kaerns-Sayre syndrome: eye defects, abnormal heartbeat, and central nervous degradation from large deletions in mitochondrial DNA.
2.3.1.5 Three-person babies
In recent times, mitochondrial DNA transfer (i.e., a three-person baby) has been seen as a way to treat mitochondrial DNA diseases. The UK is the first country to implement such laws allowing for the aforementioned.
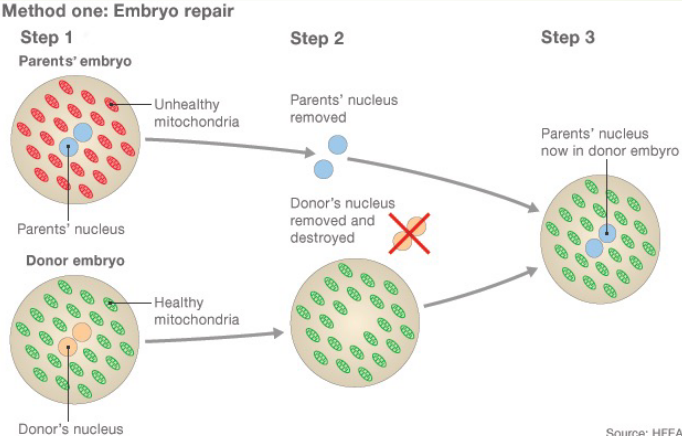
Figure 2.18: Three-Person Baby Formation Schematic
In this first method of treatment, two eggs are fertilized with sperm, creating an embryo from the parents and the donor.
The parents’ (healthy) nuclei are then removed and kept; the donor’s is destroyed and the parents’ nuclei are inserted into the donor’s, hence creating a healthy embryo.
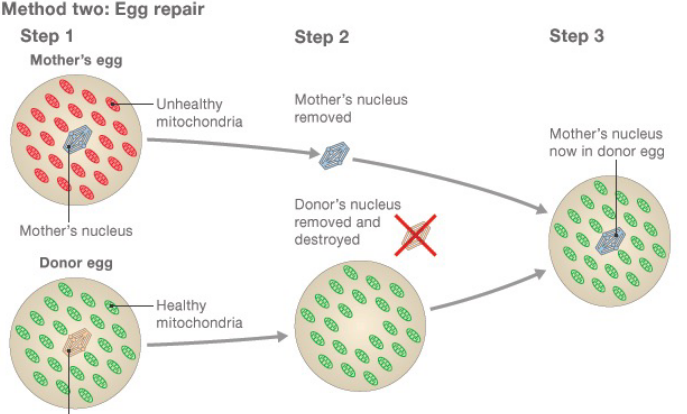
Figure 2.19: Egg Treatment in Three-Person Baby Formation
First, the mother’s (damaged) egg and the donor’s egg are collected. Then, the genetic material is removed from both eggs - the mother’s genetic material is then inserted into the donor egg’s (which can be fertilized to yield a healthy embryo).
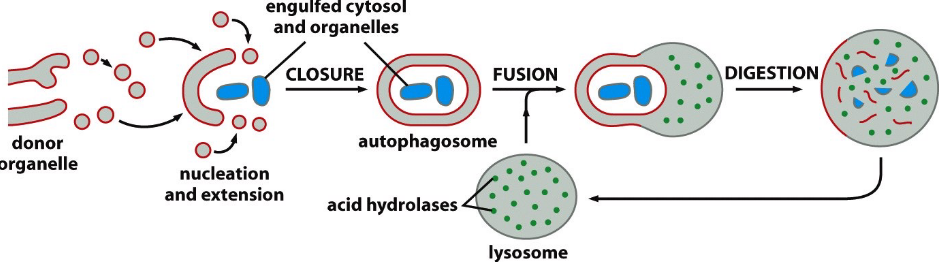
Figure 2.20: Schematic of Autophagy
Mitochondria numbers are controlled by autophagy: the process begins by wrapping a double membrane around the mitochondrion to become an autophagosome.
2.4 Plastid Genomes
To date, all plastids that are studied have their own DNA: a reduced “genome” from a cyanobacterial ancestor from the early days of the eukaryotic cell.
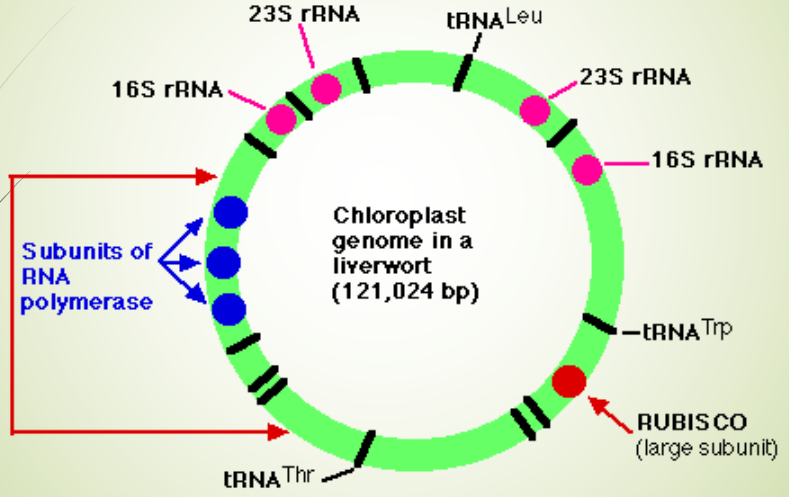
Figure 2.21: Genetic Map of a Plastid Genome
A terrestrial plant chloroplast’s genomes has about 110 - 120 unique genes. Some algae have a large chloroplast genome with more than 200 genes, while palstid genomes from non-photosynthetic organisms may have a few dozen genes.
2.4.1 Features of chloroplast genomes
Chloroplast DNA is around 70000 to 200000 base pairs long; they encode for 120 genes: 60 of which are involved in RNA transcription and translation (including rRNAs, tRNAs, and RNA polymerase subunits), 40 for proteins of unknown functions, and 20 for the F0F1 subunits of ATPase and electron-transfer complexes.
Funnily enough, chloroplasts are not inherited from male parents. Instead, it is inherited from the mother in most plants (i.e., about \(\frac{2}{3}\) of higher plants). Hence, only pollen grains contribute to the genetic information contained in the nucleus, but not in the chloroplast.
2.4.2 Origins of mitochondria and chloroplasts
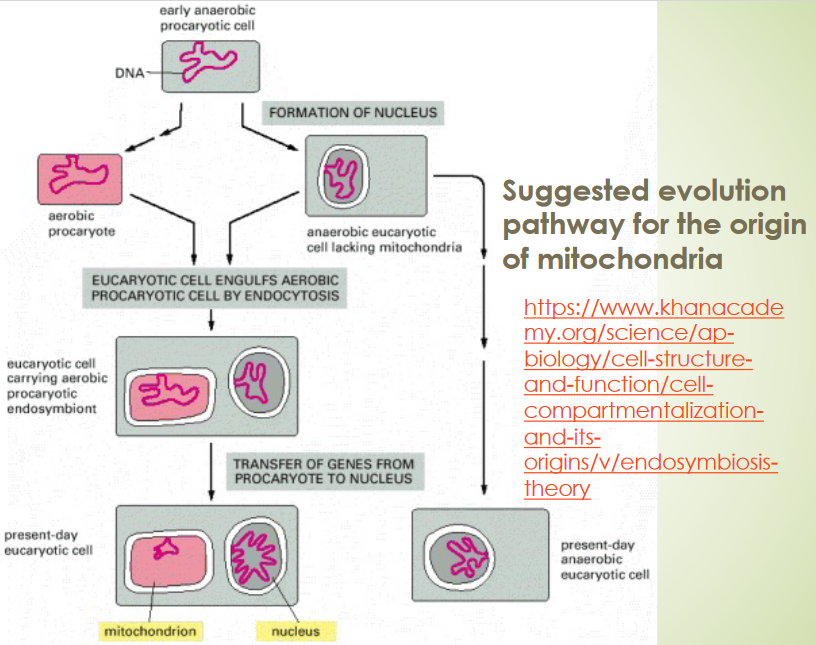
Figure 2.22: Origin of Mitochondria
The endosymbiotic theory states that:
- The mitochondria of eukaryotes evolved from aerobic bacteria living within their host cells.
- The chloroplasts of eukaryotes evolved from endosymbiotic cyanobacteria.
2.4.3 Evidence for mitochondria and chloroplasts coming from prokaryotes
Both mitochondria and chloroplasts can only form from pre-existing mitochondria and chloroplasts. They cannot be formed from a cell missing the aforementioned - nuclear genes only encode for some of the protein required by the two organelles.
Both organelles also have their own genomes.
2.4.3.1 Evidence #1
Both organelles have their own protein-synthesizing machinery - something that is more closely reminiscent of those found in prokaryotes.
2.4.3.2 Evidence #2
Both organelles’ first amino acid transcripts always begin with formyl-methionine (i.e., formyl-met).
A number of antibiotics (e.g., rifampicin) also inhibit protein synthesis in bacteria within both organelles.
2.4.3.3 Evidence #3
Inhibitors of eukaryotic ribosomes do not affect protein synthesis in both organelles.
2.5 Rickettsia and Cyanobacteria
2.5.1 Rickettsia
These are bacteria that are too small to be seen under a traditional light microscope. Almost all rickettsia are obligate, intracellular parasites. The bacterium Rickettsia prowazekii causes typhus fever when it is transmitted to humans by lice.
Rickettsia genomes can be considered the closest relatives to the ancestors of the mitochondria.
2.5.2 Cyanobacteria
These are photosynthetic prokaryotes that use chlorophyll a and water as a source of electrons to reduce CO2 to carbohydrates.
2.6 Applications of Chloroplast and Mitochondrial DNA
2.6.1 Chloroplast genetic engineering
When a foreign gene is introduced into the chloroplast’s genome, a very high transgene expression is observed.
Strangely enough, pollen does not contribute to chloroplast DNA in the zygote - the risk of transgene escape is rare if the gene of interest is inserted into the chloroplast genome.
2.6.1.1 Application #1: glyphosphate resistance
Glyphosphate is a potent, broad spectrum herbicide that is effecting in suppressing the growth of most grasses and broadleaf weeds.
This chemical affects the biosynthesis of aromatic amino acids in plants and microorganisms - it is not toxic to animals.
In 1998, Daniell et al. successfully transformed a gene conferring glyphosphate resistance into the tobacco chloroplast genome (hence, this resistance gene cannot be transferred to other plants via pollination); the transgenic plants were able to survive in environments that have ten times the lethal concentration of glyphosphate.
Some examples of glyphosphate-resistant plants include: soy, corn, alfalfa, sugar beets, and cotton.
2.6.1.2 Application #2: insect resistance via Bt toxin
Bacillus thuringiensis (i.e., Bt) toxin is harmful to insects when ingested.
In 1999, Kota et al. saw the hyper-expression of Bt toxins in transformed tobacco chloroplasts that led to a high mortality rate in insects.
The toxins in such an application are located in the green, leafy tissues (i.e., where insects are most likely to eat the plant) for the most part.
2.6.2 Evolutionary trees
Mitochondrial DNA is used for this purpose; this is done by measuring the substitution rate of mitochondrial genomes.
2.6.2.1 Why mitochondrial DNA?
Mitochondrial DNA are present in large numbers in each cell, so fewer samples are required. Such DNA also have a higher rate of substitution than nuclear DNA, hence making it easier to resolve differences between closely-related individuals.
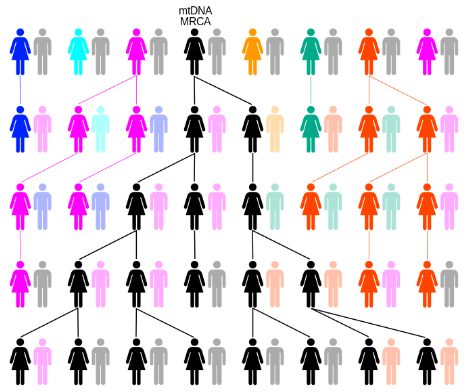
Figure 2.23: Tracing Ancestors Using Mitochondrial DNA
Furthermore, mitochondrial DNA do not recombine and are only inherited from the mother (hence allowing for the tracing of a direct genetic line).
2.6.3 Additional roles of mitochondrial DNA
Mitochondria play an important role in apoptosis.
2.6.3.1 Bcl-2 protein family
bcl-2 proteins are a protein family that is involved in apoptosis response. Some of these proteins (e.g., bcl-2 and bcl-XL) are anti-apoptotic, while some of these proteins (e.g., Bad and Bax) are pro-apoptotic. Cells are more sensitive to apoptosis when there is an excess of pro-apoptotic proteins and vice versa.
Pro-apoptotic proteins are found in the cytoplasm and act as sensors of cellular damage or stress. During periods of cellular stress, they relocate to the surface of the mitochondria (where anti-apoptotic proteins are found).
The above interaction of anti and pro-apoptic proteins creates pores in the mitochondria and release cytochrome C and other pro-apoptotic molecules from the intermembrane space.
The above leads to the formation of the apoptosome and the activation of the caspase cascade.
2.6.3.2 Caspase cascade
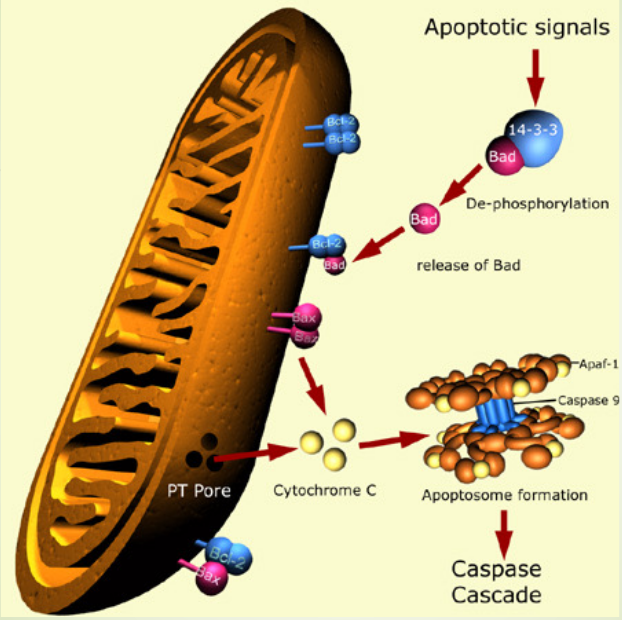
Figure 2.24: Diagram of the Caspase Cascade
When cytochrome C is released from the mitochondria, the protein caspase 9 and subsequently, caspase 3 can be activated. This effect is mediated through the formation of an apoptosome: a multi-protein complex that has cytochrome C, Apaf-1, pro-caspase 9, and ATP.