Topic 6 Cell Communication
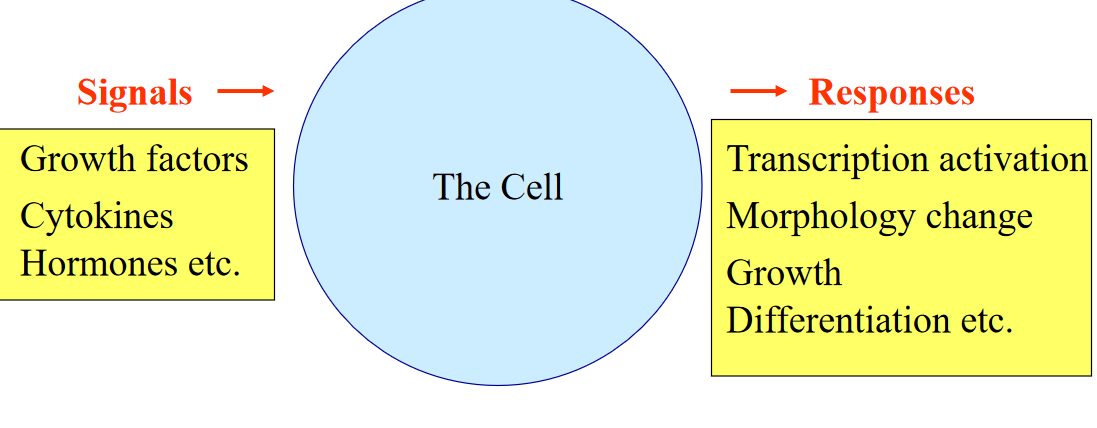
Figure 6.1: A Flowchart of Signal Transduction
The above diagram illustrates the basic idea behind cellular communications.
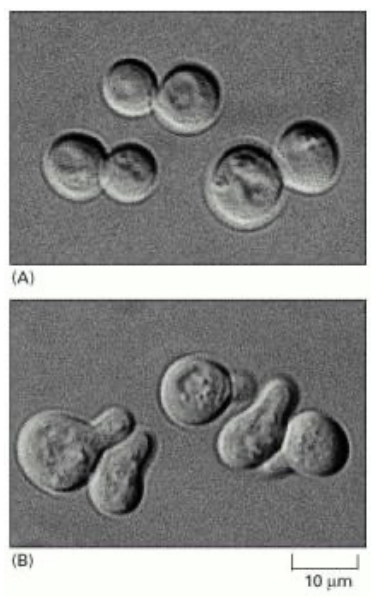
Figure 6.2: Yeast Cells Budding in Response to a Mating Factor
The above image shows a handful of yeast cells budding in response to a mating factor.
Nonetheless, different kinds of organisms require cellular communication for various purposes:
Unicellular organisms
Yeast, slime moulds, and other microbes need cellular communication to mate, aggregate, and differentiate into other cell types.
Multicellular organisms
Cellular communication is needed for the proper development and differentiation of cells and other metabolic functions.
6.1 Types of Signalling
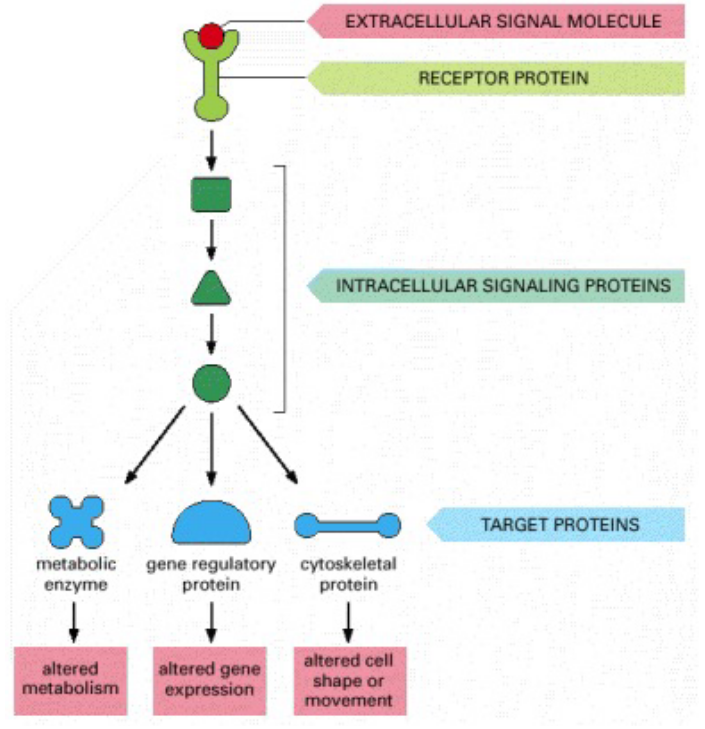
Figure 6.3: Intracellular Signalling
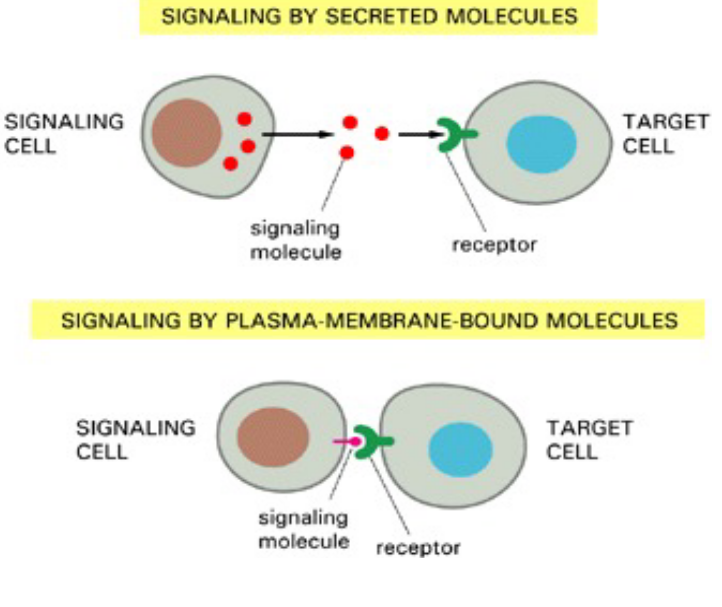
Figure 6.4: Extracellular Signalling
Note the above two signalling pathways. Furthermore, there are also many signalling molecules to note:
Amino acids
For instance, glutamate and glycine
Polypeptides
For instance, insuling and growth factors
Small peptides
Such as vasopressin and oxytocin
Nucleotides
Such as GTP and cyclic AMP
Steroids
Such as estrogen and progesterone
Retinoids
For instance, retinoic acid
Fatty acid derivatives
Such as arachidonic acid
Gases
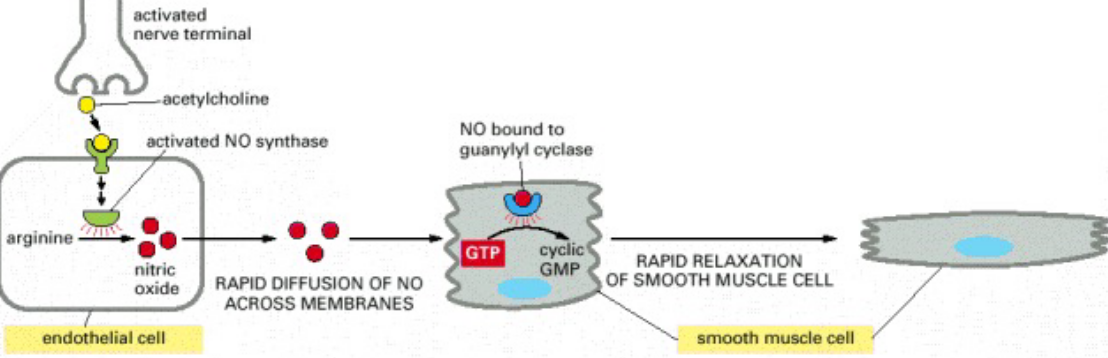
Figure 6.5: NO in Smooth Muscle Relaxation
Nitroglycerine has been used for about a 100 years to treat patients with angina. When nitroglycerine enters the body, it is converted into NO gas - this gas relaxes blood vessels, hence also reducing the workload on the heart and also its heart muscles.
Most water-soluble molecules are broken-down within minutes of entering the organism’s bloodstream - local mediators and neurotransmitters are removed even faster (i.e., within miliseconds and seconds)!
Steroid hormones can persist for days!
For this reason, water-soluble molecules are generally used for responses with short durations and water-insoluble molecules for responses with longer durations.
6.1.1 Intracellular signalling
There are four main types of intracellular signalling covered in BS2004:
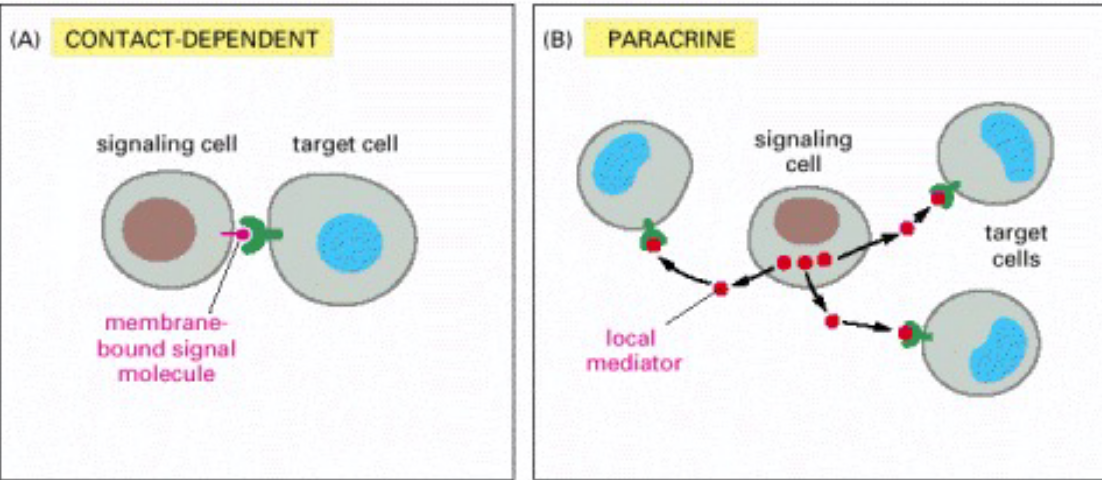
Figure 6.6: Types of Intracellular Signalling
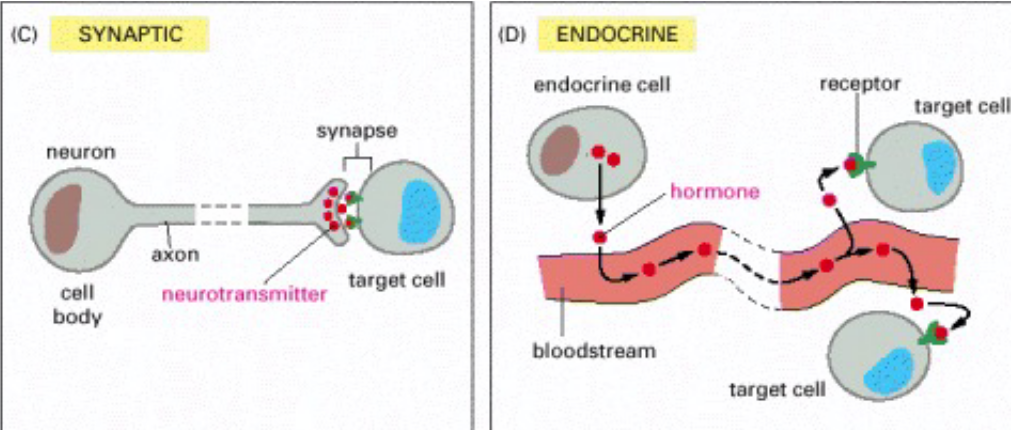
Figure 6.7: More Types of Intracellular Signalling
Contact-dependent
Paracrine
Synaptic
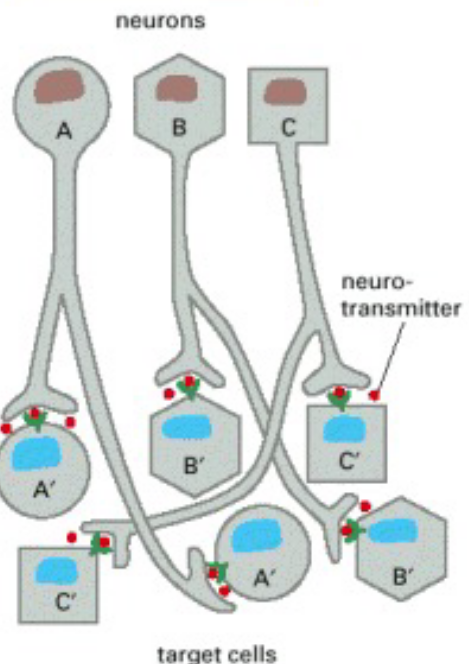
Figure 6.8: Synaptic Signalling Illustrated
The specificity of this kind of signalling arises from the contact between the nerve cell and its target cells.
Endocrine
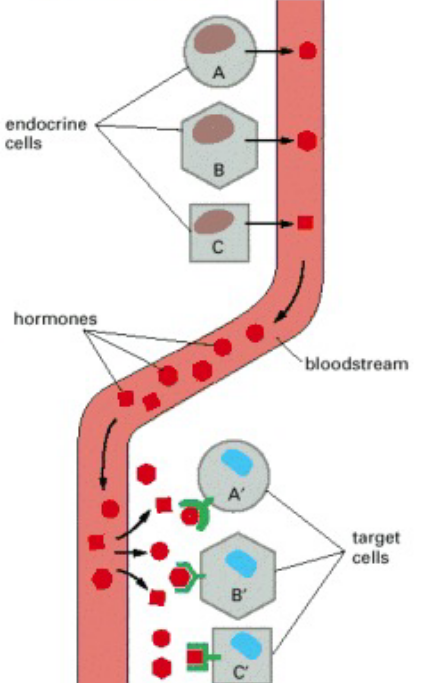
Figure 6.9: Endocrine Signalling Illustrated
In this kind of intracellular signalling, endocrine cells secrete signal hormones - hormones - into the bloodstream.
As seen above, the target cells may be very far away from the endocrine cells themselves; the target cells also have receptors that are able to bind to hormones.
Autocrine
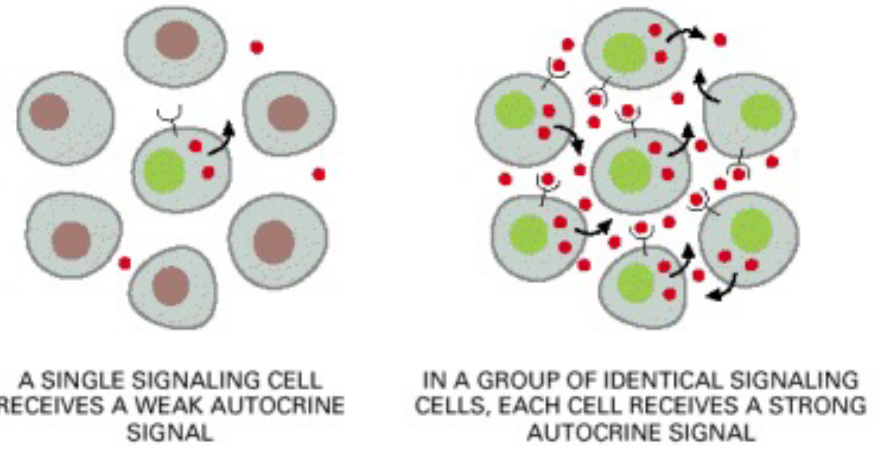
Figure 6.10: Endocrine Signalling Illustrated
In this kind of intracellular signalling, cells release autocrine signals that act on their own kind.
Cancer cells often use this kind of signalling to overcome cell proliferation and survival.
6.1.1.1 Differences between endocrine and synaptic signalling
In long range signalling, the target cell can be very far away. Hence, in terms of…
Speed
Synaptic signalling is faster and more precise. Endocrine signalling is slow because secreted hormones need time to reach the target cell via the bloodstream.
Concentrations
Neurotransmitters work at higher concentrations (e.g., 5 \(\times\) 10-4 M for acetylcholine). Hormones don’t require that high a concentration (typically less than 10^-8$ M).
6.2 (Gap) Junctions
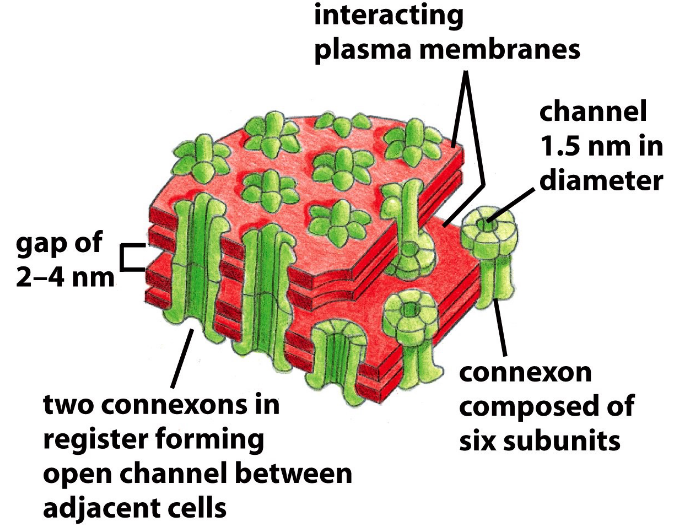
Figure 6.11: A Schematic of Gap Junctions
Gap junctions are an aggregate of channels that join adjacent cells. Intracellular channels are made up of a pair of connexons - a hexamer of connexin proteins.
Nevertheless, gap junctions also help coordinate the activity of neighboring cells - for instance, by allowing intracellular signalling molecules such as Ca2+ and cAMP to be exchanged.
6.2.1 Different cells, different responses
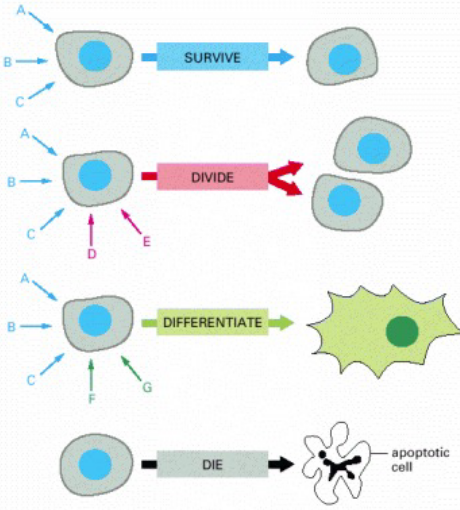
Figure 6.12: Possible Reactions of Cells to Signals
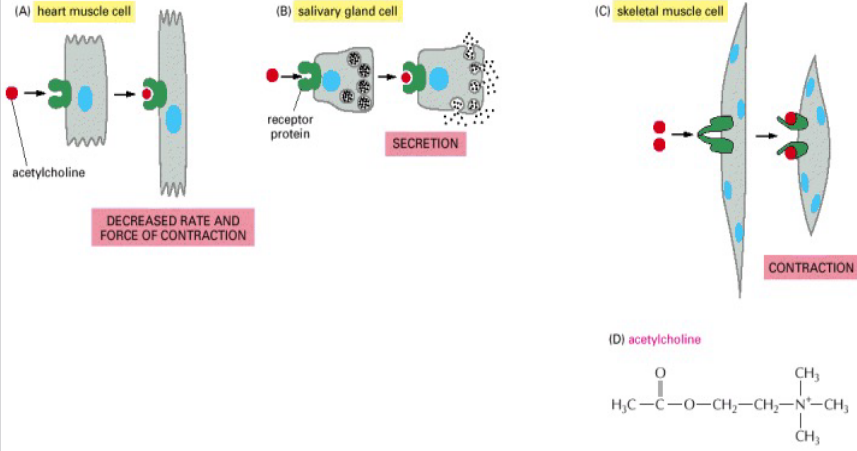
Figure 6.13: Acetylcholine Causing Different Reactions in Different Cells
As seen above, the neurotransmitter acetylcholine can have different effects on different cells - this is because of:
Different receptors
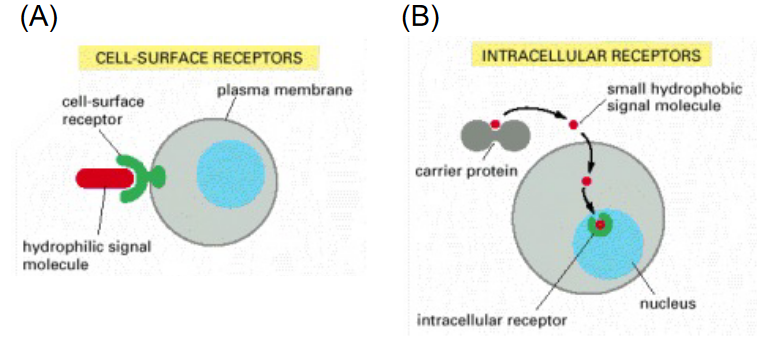
Figure 6.14: Different Kinds of Receptors
Different internal signalling molecules
Different target genes or activated genes
6.3 Intracellular Receptors
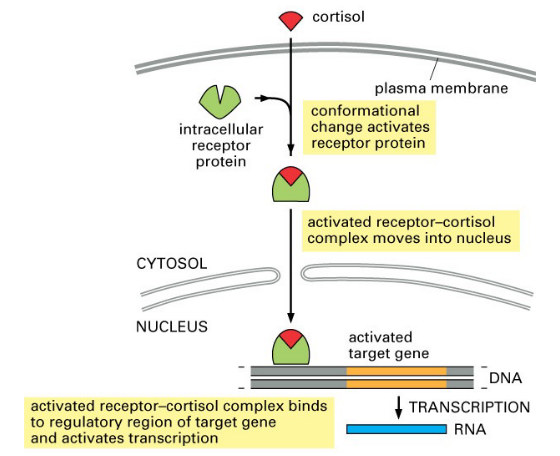
Figure 6.15: Example Mechanism of Cortisol Interacting with an Intracellular Receptor Protein
A small number of hydrophobic signals diffuse across the plasma membrane of target cells and bind to intracellular receptor proteins. These signals can include thyroid hormones, retinoids, and vitamin D.
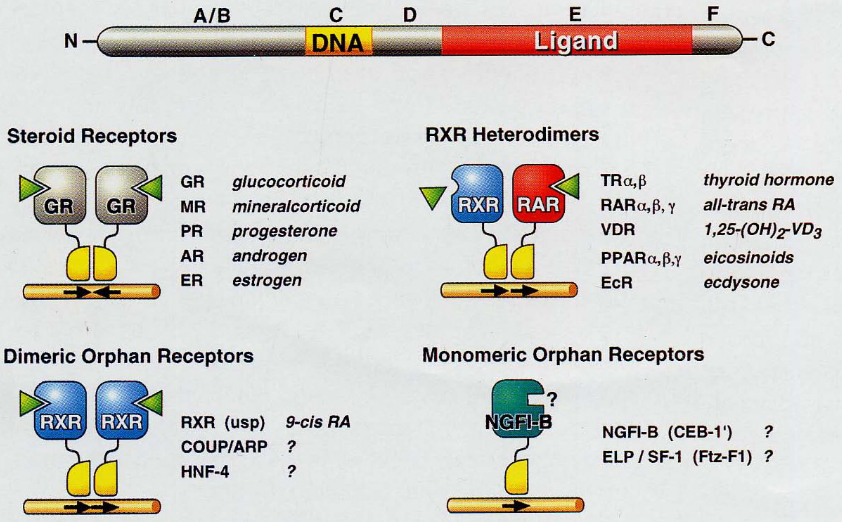
Figure 6.16: Different Kinds of Nuclear Receptors
Most times, the receptor themselves belong to the nuclear receptor superfamily.
6.3.1 Type I and II receptors
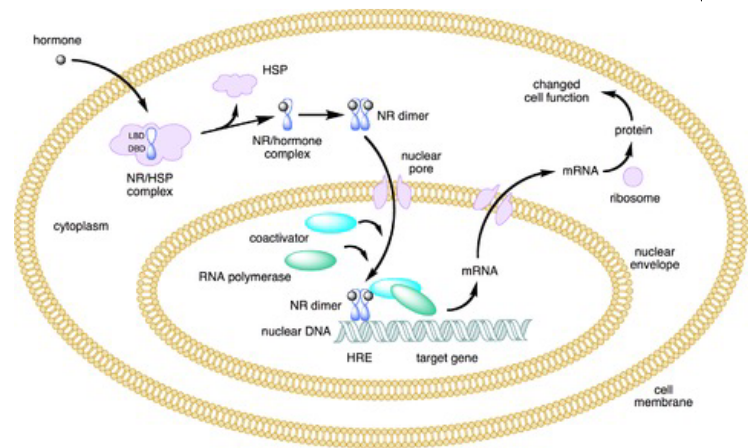
Figure 6.17: Type I Nuclear Receptors
Type I receptors have a heat shock protein (i.e., hsp) associated with inactive receptors that will be released when the receptor interacts with a ligand. The receptors then dimerizes and translocates to the nucleus. Once inside the nucleus, the receptors then bind to specific DNA sequences to influence transcription.
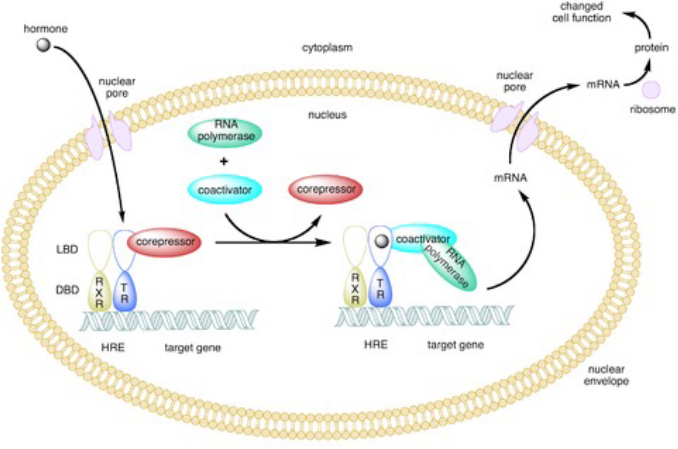
Figure 6.18: Type II Nuclear Receptors
Type II receptors lack a hsp and are found in the cell’s nucleus. Inhibitory proteins may bind to type II receptors in the nucleus to keep them in their inactive form. Ligand binding to these receptors causes these inhibitory proteins to disassociate and recruit transcription co-factors.
6.3.2 Surface receptors
The three largest classes of surface receptors are the:
- Ion channel-coupled receptors
- G-protein coupled receptors
- Enzyme-coupled receptors
6.3.2.1 Ion channel-linked receptors
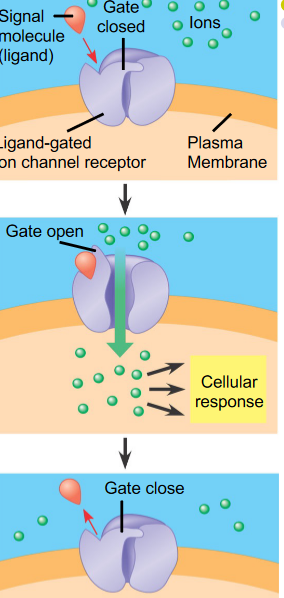
Figure 6.19: Ion Channel-Linked Receptors
For instance, neurotransmitters that open or close an ion channel formed by the protein to which they bind to, hence changing the ion permeability of the plasma membrane and the excitability of the post-synaptic cell.
Multipass transmembrane proteins are a good example of this!
6.3.2.2 Enzyme-coupled receptors
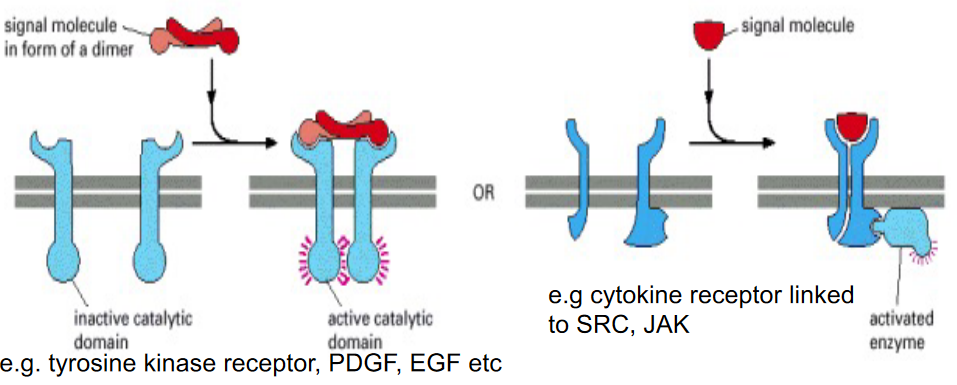
Figure 6.20: Enzyme-Coupled Receptors
These either function as enzymes or are associated with the enzymes that they activate.
These kind of receptors are single-pass transmembrane proteins with their ligand-binding sites on the outside of the cell and their catalytic or enzyme-binding site on the inside.
6.3.2.3 G-protein coupled receptors
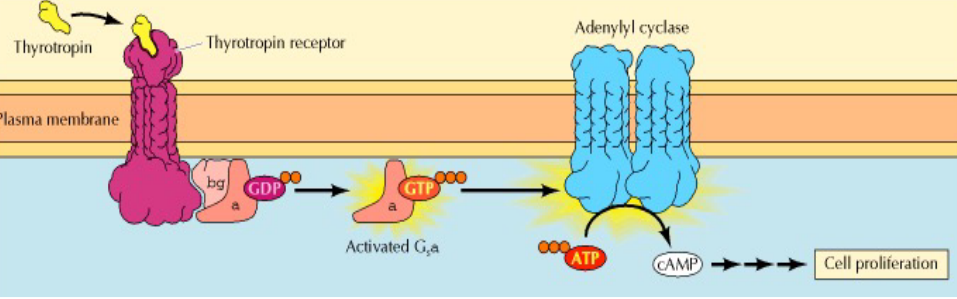
Figure 6.21: G-Protein Coupled Receptors
These receptors indirectly regulate the activity of a separate plasma-membrane bound target protein (which can either be an enzyme or an ion channel).
A trimeric GTP-binding protein (i.e., G protein) mediates interactions between these receptors and the target protein.
These receptors are very widespread and have diverse functions:
- Embryonic development
- Vision and hearing
- Yeast mating
- Hormone systems
- Neurotransmitter signalling pathways
For this reason, it intuitively makes sense that faulty G-protein coupled receptors are associated with many diseases (e.g., cholera, botulism, etc). 60% of all medicine to date also have some sort of effect on the performance of G-protein pathways.
6.3.3 More on G proteins and signalling
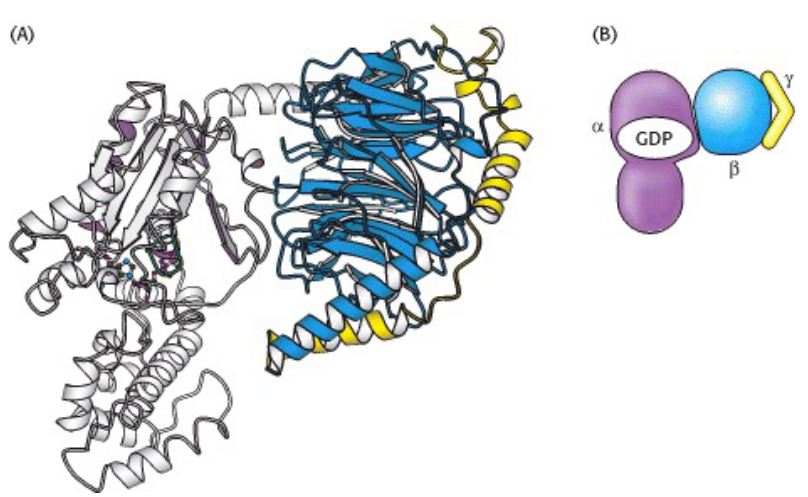
Figure 6.22: Structure of a Heterotirmeric G Protein
Heterotirmeric G proteins are linked to a G protein-coupled receptor: also known as a 7-transmembrane receptor.
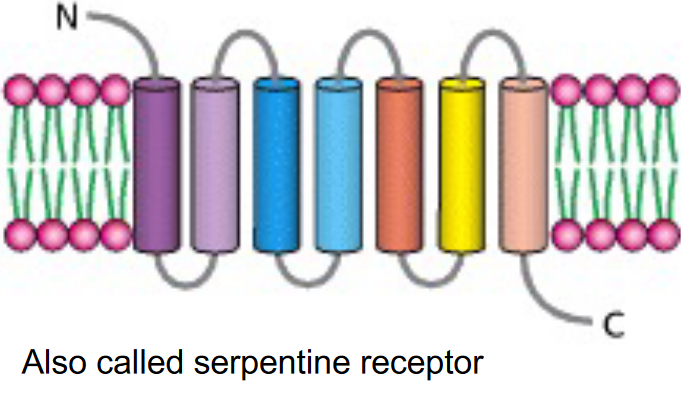
Figure 6.23: A 7-Transmembrane Receptor
These 7-transmembrane receptors have three subunits: \(\alpha\), \(\beta\), and \(\gamma\).
6.3.3.1 Mechanism of action for trimeric G proteins
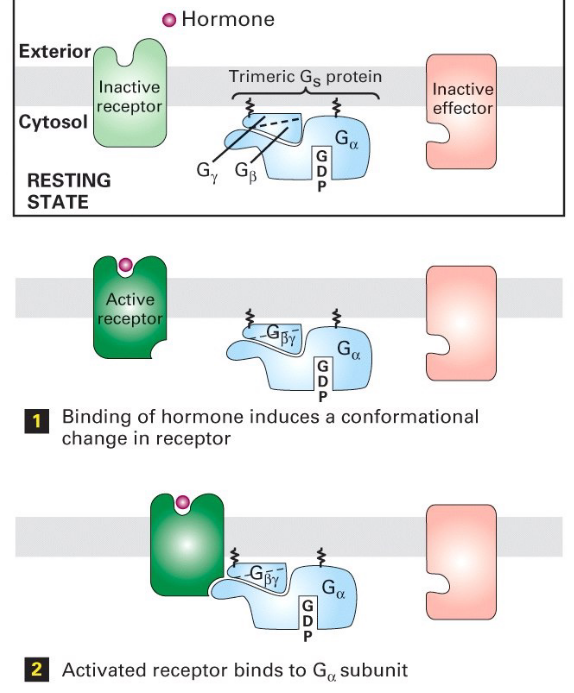
Figure 6.24: Steps 1 - 3 of Trimeric G Protein Activation
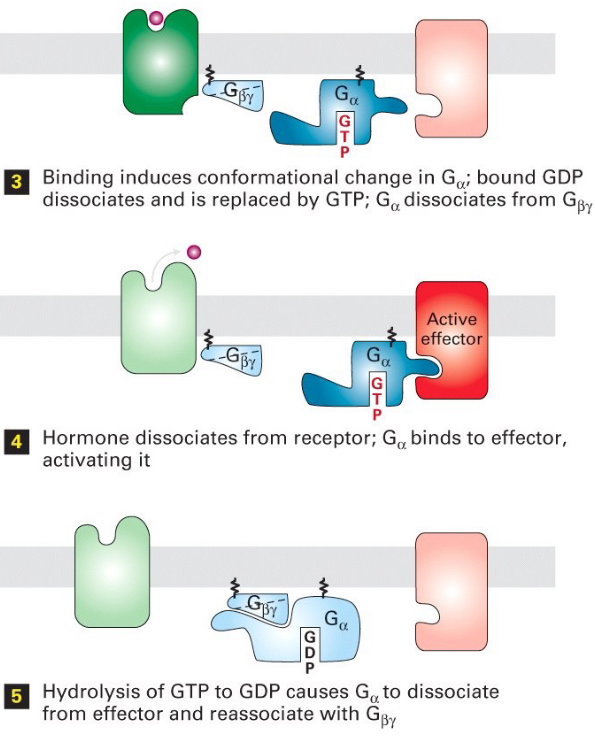
Figure 6.25: Steps 4 - 6 of Trimeric G Protein Activation and Deactivation
Note the following steps:
- G-protein (with GDP) loosely binds to the receptor
- Signal molecule binds the receptor
- Binding induces a conformational change in the receptor
- Conformation change induces the receptor to activate the G-protein
- \(\alpha\) subunit of the G-protein releases GDP and binds GTP
- The conformational change on G\(\alpha\) (i.e., the \(\alpha\) subunit) then exposes the \(\beta\) and the \(\gamma\) subunits to allow interactions with their target proteins.
- The \(\alpha\) subunit becomes a GTPase - when GTP becomes hydrolyzed to GDP, the G protein becomes inactive.
6.3.3.2 Signal transduction
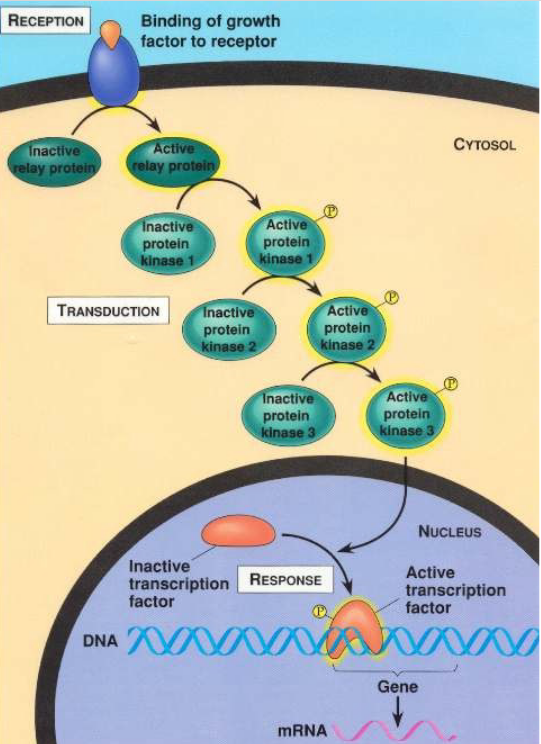
Figure 6.26: An Example of Signal Transduction
Most activated cell surface receptors relay their signals via small molecules and a network of intracellular signalling proteins.
6.3.3.3 Second messengers
These are a diverse family of low molecular weight compounds (e.g., cAMP and Ca2+ ions) that transmit biological signals initiated by receptor-ligand binding at the cell surface to intracellular targets or effector proteins.
Some types of second messengers include:
Hydrophobic molecules
For instance, diacylglycerol and phosphatidylinositols - all of these are membrane-associated and regulate membrane-associated effector proteins.
Hydrophilic molecules
For instance, cAMP, cGMP, and Ca2+ ions - these are all found within the cytosol.
Gases
For instance, NO gas and CO gas - these can diffuse through the cytosol and cellular membranes.
6.3.3.3.1 Generations of second messengers
Adenylyl cyclase
This is an enzyme that catalyzes the formation of cAMP from ATP. This enzyme is also called 3’,5’-cyclic AMP synthetase.
Phospholipase C
This is an enzyme that catalyzes the formation of inositol-1,4,5-triphosphate (i.e., IP3) and diacylglycerol from phosphatidylinositol-4,5-biphosphate (i.e., PIP2).
6.3.3.3.2 cAMP
Cyclic AMP (i.e., cAMP) is a small intracellular mediator with roles in prokaryotic and animal cells.
cAMP exists in cells at concentrations of around 10-7 M, but extracellular signals can increase their levels by as much as 20 fold in 20 seconds. This rapid response needs some sort of rapid synthesis mechanism and a rapid removal of this mediator!
cAMP is made from ATP via a membrane-bound enzyme called adenylyl cyclase - cAMP is continuously destroyed by cAMP phosphodiesterases that hydrolyze cAMP to adenosine monophosphate (i.e., 5’-AMP)