3.8.1.2 Primary cilium
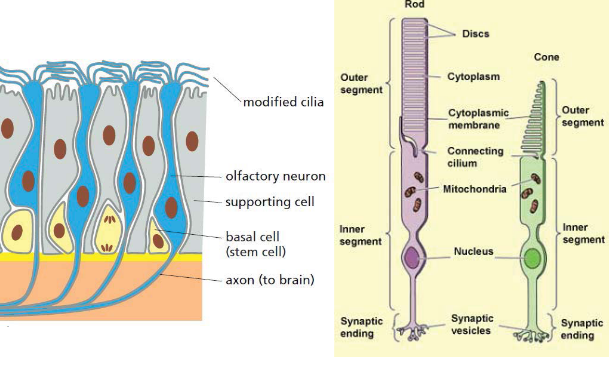
Figure 3.24: Short Cilia
These are used for sensing inputs - almost all cells in the human body have cilia.
Eukaryotic cells have many organelles compared to prokaryotes. An organelle is a sub-cellular membrane-enclosed compartment that has a distinct structure, composition, and function.
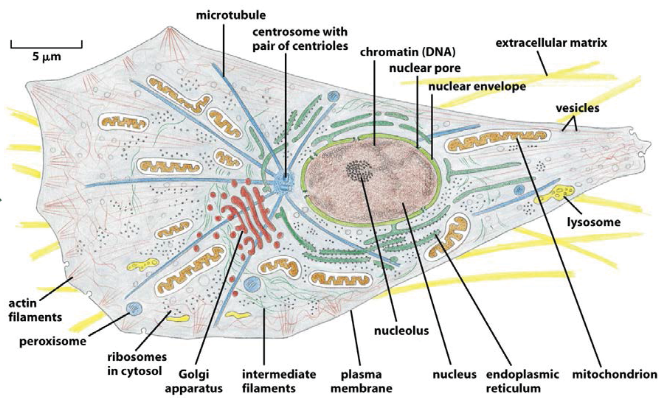
Figure 3.1: Some Eukaryotic Organelles
Topology is the study of shapes and spaces.
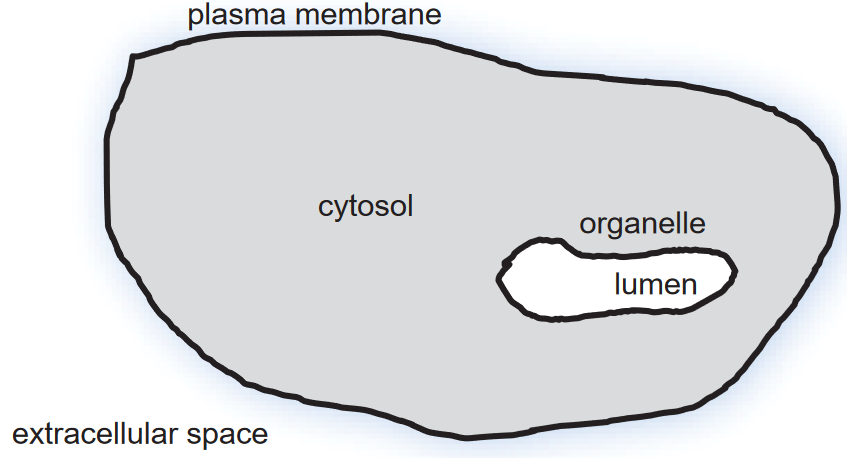
Figure 3.2: Topology of a Cell
Organelles are typically formed by closed lipid membranes.
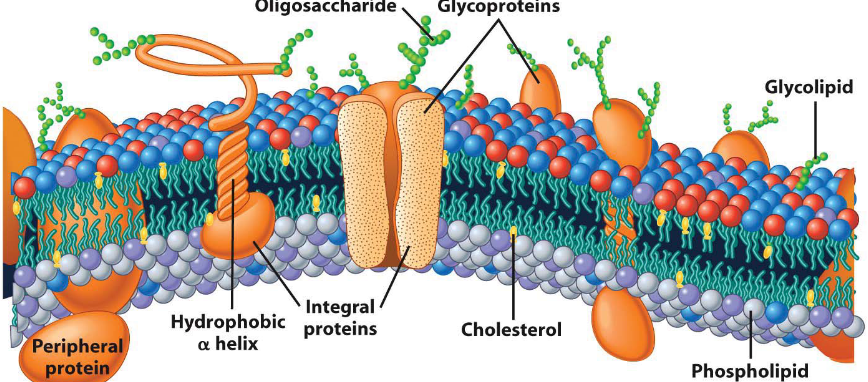
Figure 3.3: Textbook Depiction of a Biological Membrane
There are three main components to a biological membrane:
Lipids
These account for about 50% of the mass of the membrane. Lipids also serve as a barrier to prevent the movement of hydrophilic materials in and out of organelles.
Proteins
These also account for about 50% of the membrane’s mass. Proteins contribute to the membrane’s function.
Carbohydrates
Several carbohydrate groups are present in figure 3.3.
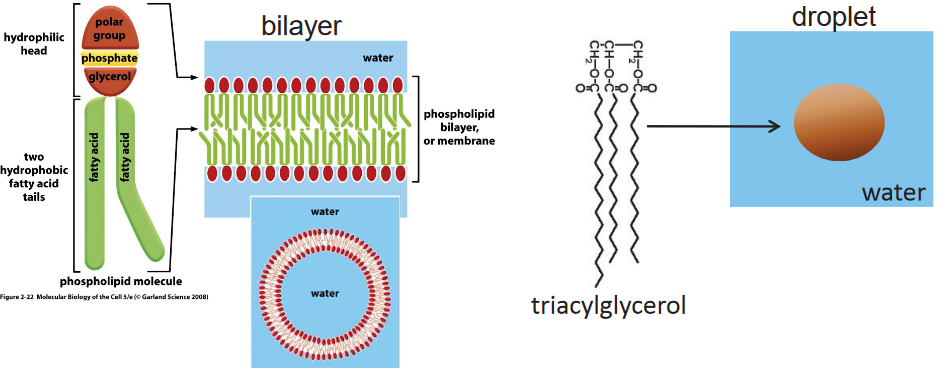
Figure 3.4: Aggregation of Membrane Lipids and a Single Triacylglycerol Molecule
Membrane lipids can be classified into one of three categories:
All membrane lipids have a hydrophobic tail and a hydrophilic head. Because of this, we say that membrane lipids are amphipathic: they have a polar and a non-polar region.
Like figure 3.4 suggests, membrane lipids are capable of self-assembly and can form a lipid bilayer in aqueous environments.
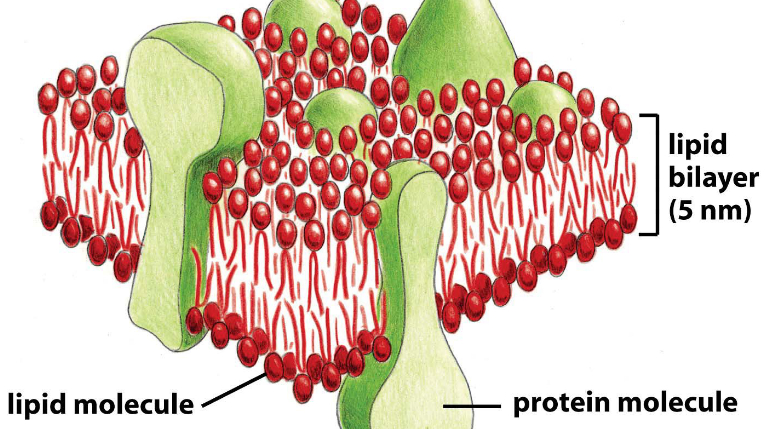
Figure 3.5: A Leaflet of the Lipid Bilayer
A leaflet is a unit of the cytoplasmic and the extracellular membrane. It is also the universal feature of biological membranes.
All leaflets are impermeable to hydrophilic molecules are fluid in two dimensions: a leaflet cannot flip-flop!
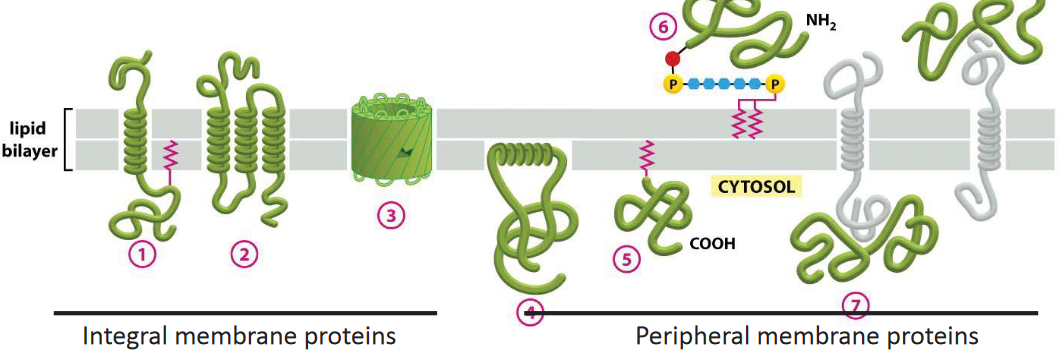
Figure 3.6: Several Membrane Proteins
There are two main classes of membrane proteins in figure 3.6:
Integral membrane proteins
Such membrane proteins can be predicted from looking at their primary structures. A transmembrane domain is about 20 amino acids long and generally form an \(\alpha\)-helix.
Peripheral membrane proteins
These attach to integral membrane proteins or lipids and cycle between the cytosol and the membrane.
There are distinct biochemical properties between integral and peripheral membrane proteins: peripheral membrane proteins are more easily disassociated under harsh chemical conditions (e.g., high salinity).
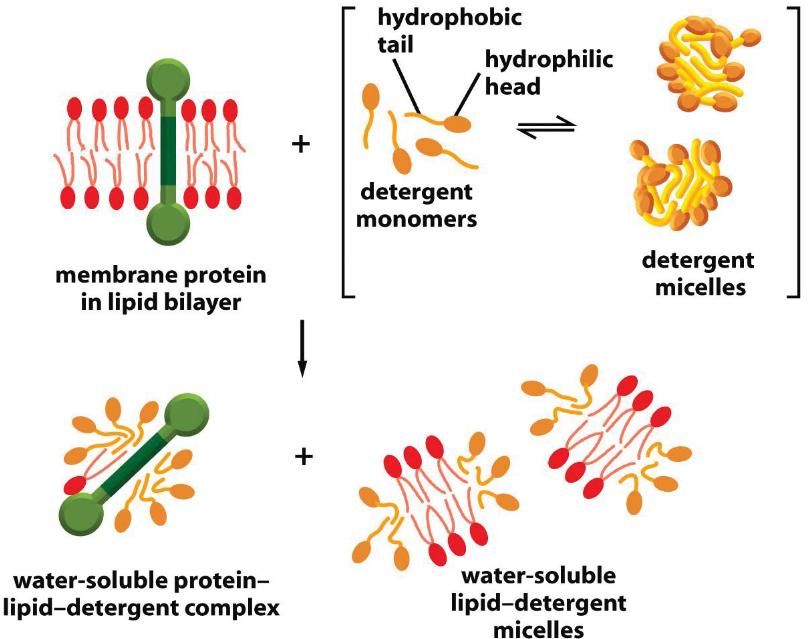
Figure 3.7: Membrane Solubilizing with Detergents
Observe the above graphic.
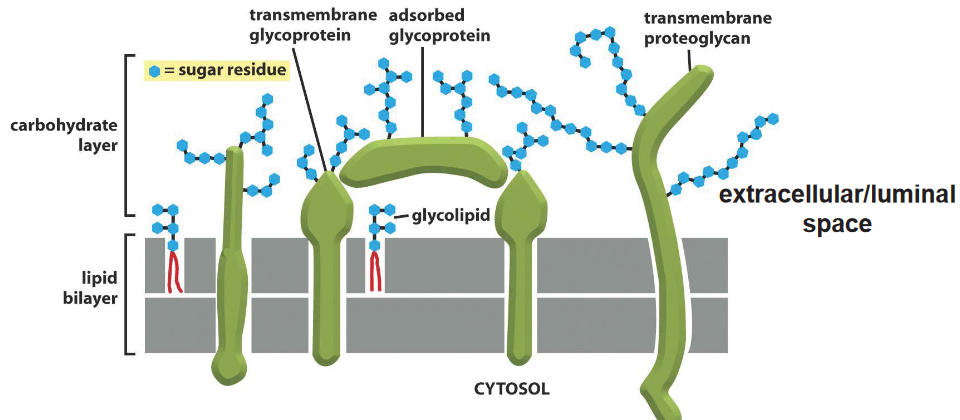
Figure 3.8: Several Membrane Carbohydrates
Figure ?? above shows several carbohydrates that are attached to a protein or a lipid. When attached to either one of the aforementioned structures, they are called glycoproteins or glycolipids.
Membrane carbohyrates are also randomly located in the extracellular or the luminal space.
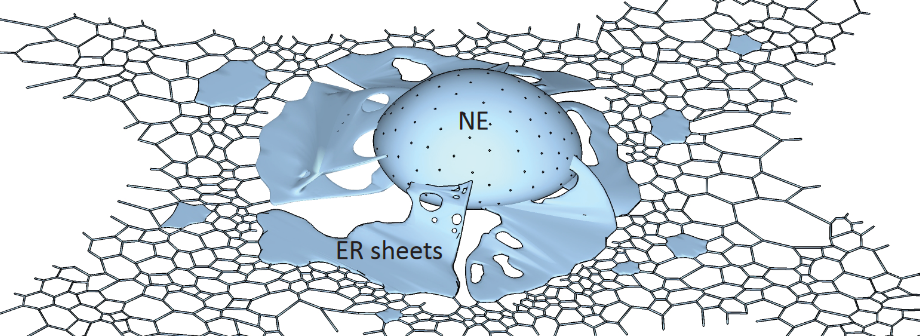
Figure 3.9: Schematic of the Nuclear Membrane
This is the largest organelle (i.e., takes up about 50% of the cell’s total membrane). It is made up of membrane sheets and also tubules; it also has a continuous membrane and a luminal space that also assembles the nuclear envelope.
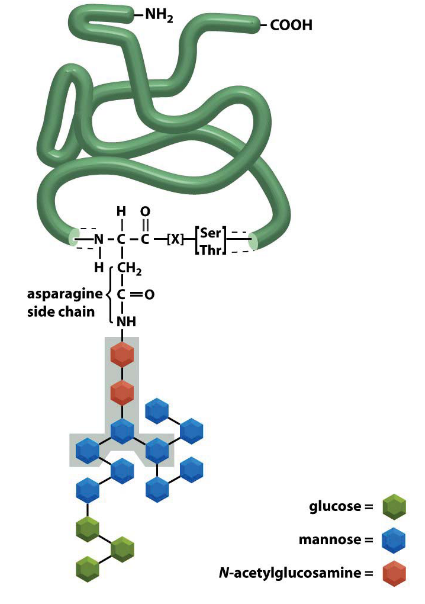
Figure 3.10: N-Glycosylation
N-Glycosylation of proteins also happen in the ER.
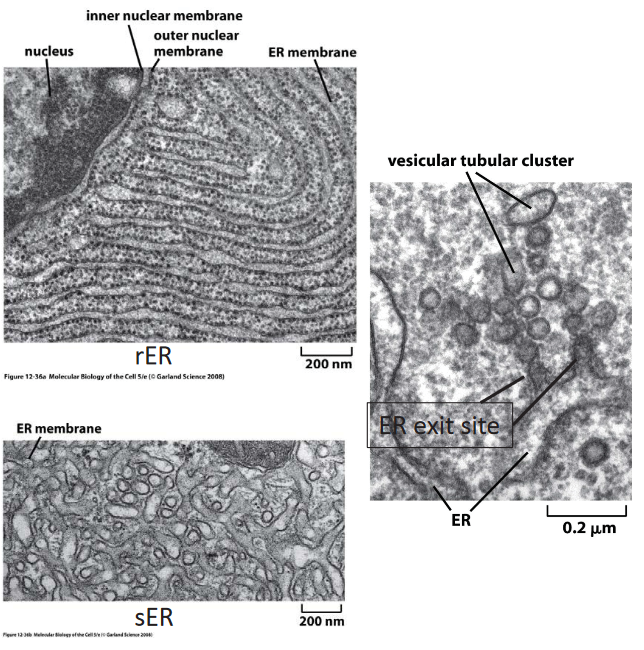
Figure 3.11: Domains of the ER
It has three main components:
Rough ER (i.e., rER)
This portion of the ER is attached to ribosomes and primarily serves to synthesize secreted proteins.
Smooth ER (i.e., sER)
This portion of the ER is devoid of ribosomes.
ER exit site
Here, COPII-coated membranes bulge out from the ER. Proteins also get secreted (i.e., exported) and recruited in this portion of the ER.
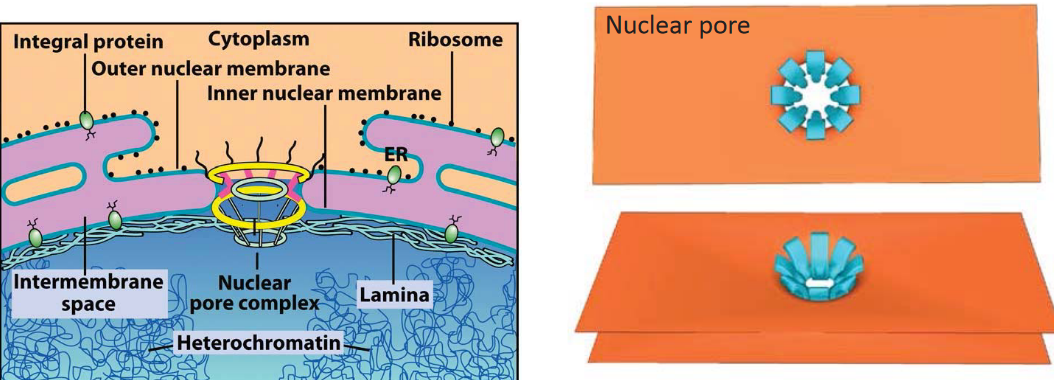
Figure 3.12: Diagram of the Nuclear Envelope
This organelle is the defining nature of eukaryotes - the nuclear membrane has two membranes.
The nuclear pore is an aqueous channel and the only type of connection between the nucleus and the cytosol. The pore itself exhibits 8-fold rotational symmetry and also facilitates nucleocytoplasmic trafficking.
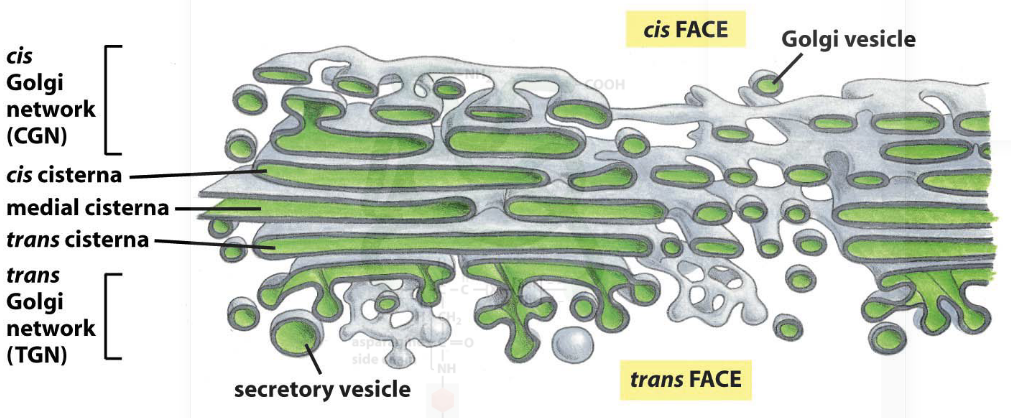
Figure 3.13: Diagram of the Golgi Complex
The golgi complex is an organelle that is made up of many stacks of laterally-connected mini stacks.
One can think of a “mini-stack* as the”functional unit" of the golgi complex. It is made up of four to seven membrane sacks called cisternae (cisterna in singular form), is polarized (note the multiple layers of the complex and how cargo enters from the cis site and leaves on the trans site).
Also note how there are many budding profiles in the golgi complex.
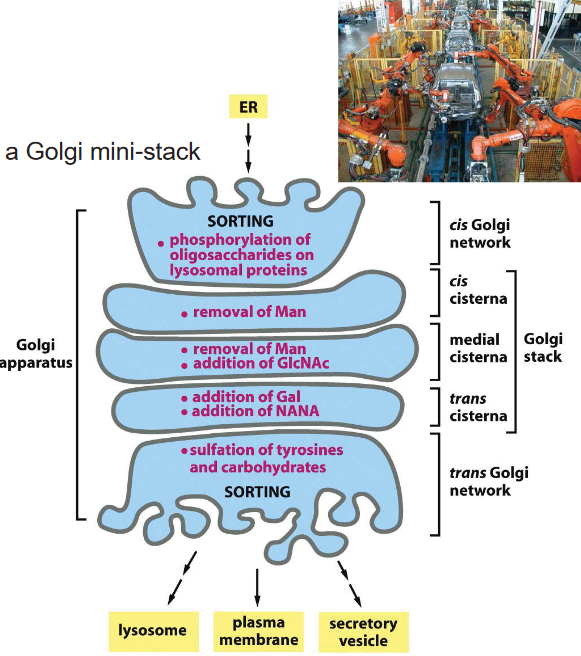
Figure 3.14: Oligosaccharide Processing in the Golgi Complex
Oligosaccharide processing in the golgi complex is sequential: it always proceeds in the cis to the trans direction. Because of this, enzymes that are involved in the early phase of oligosaccharide processing are always located in the cis end of the complex and vice versa!
The enzymes glycosidase and glycosyl transferase are resident enzymes of the golgi complex.
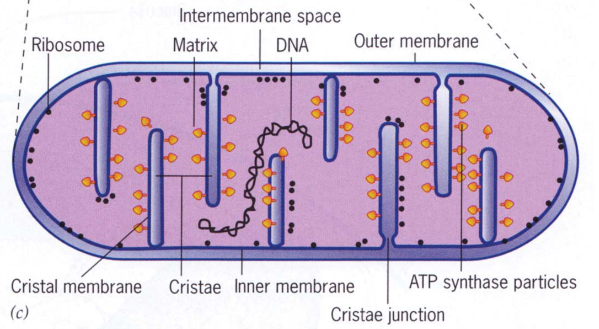
Figure 3.15: Cross-Section of a Mitochondrion
Mitochondria are organelles that are generally depicted as long rods that run along microtubules.
Mitochondria also have three different areas:
The matrix of the mitochondria has two main components: circular DNA and ribosomes.
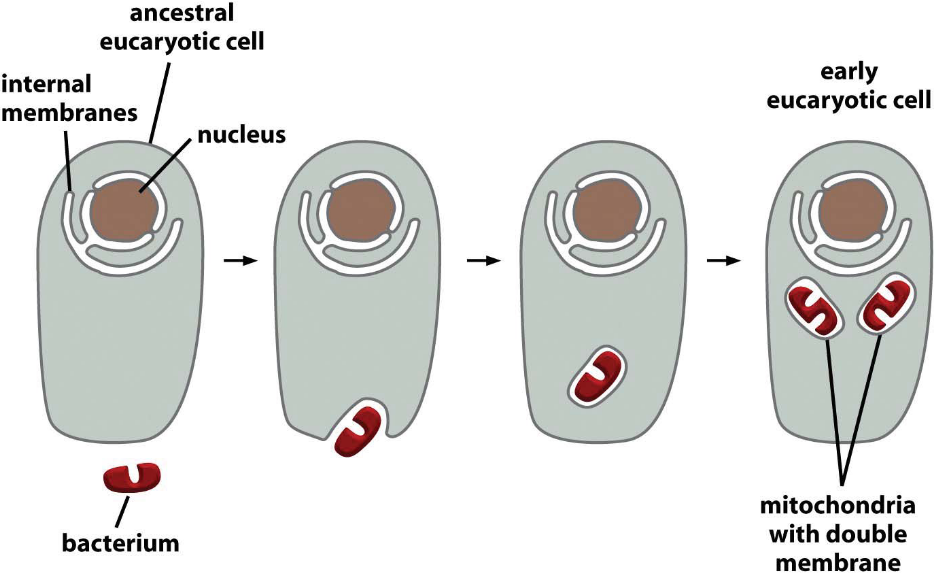
Figure 3.16: Evolution of the Mitochondria
It is common believed that mitochondria evolved via endosymbiosis of a bacterium.
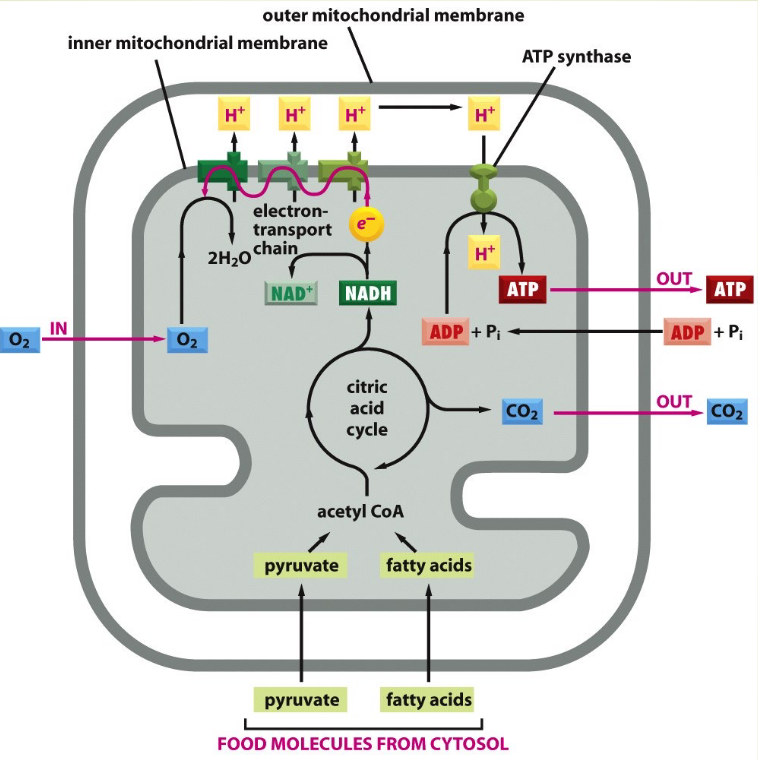
Figure 3.17: ATP Production in the Mitochondria
Here, a proton gradient is generated across the inner membrane.
The electric field strength is defined by:
\[\begin{equation*} \frac{\text{Voltage}}{\text{Distance}} \end{equation*}\]
Hence, the electric field strength of a mitochondrion is 30 million volts per meter!
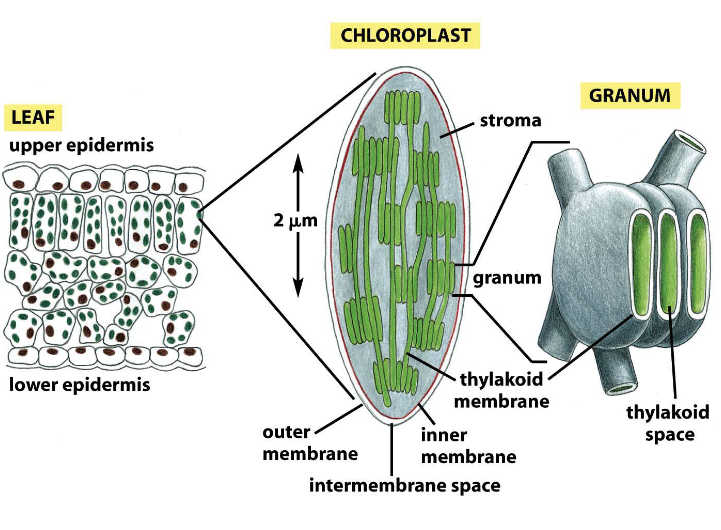
Figure 3.18: Chloroplast Structure
Like the mitochondria, the chloroplast also has an inner membrane, an outer membrane, and an inter-membrane space. However, its inner membrane lacks cristae.
Its stroma has circular DNA and ribosomes. Note how the granum is made up of stacks of thylakoids.
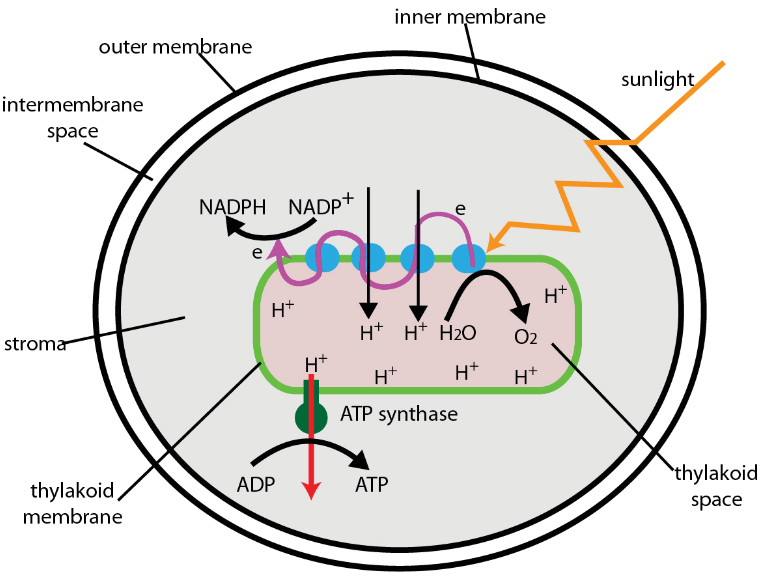
Figure 3.19: Electron Transport in the Chloroplast
The electron transport chain is located on the surface of the thylakoid membrane - the proton gradient is generated across the thylakoid membranes.
Electron transport serves to produce ATP and perform carbon fixation.
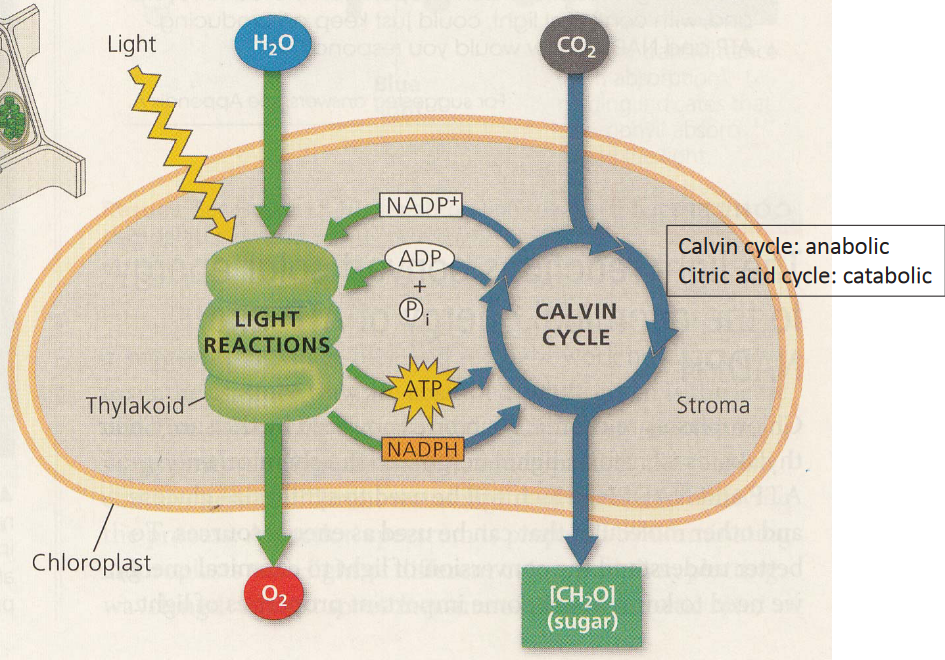
Figure 3.20: Reactions of Photosynthesis
Here, note that photosynthesis has two different reactions.
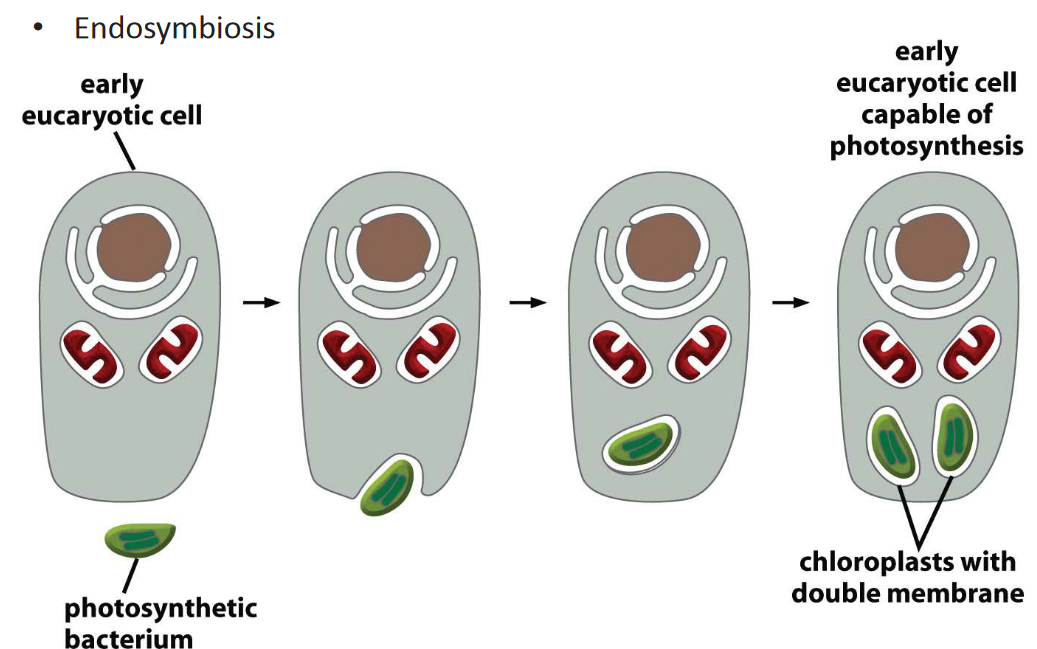
Figure 3.21: Origin of Chloroplasts
It is also common believed that chloroplasts originated via endosymbiosis.
These organelles are a protrusion of the plasma membrane - in essence, a bundle of microtubules.
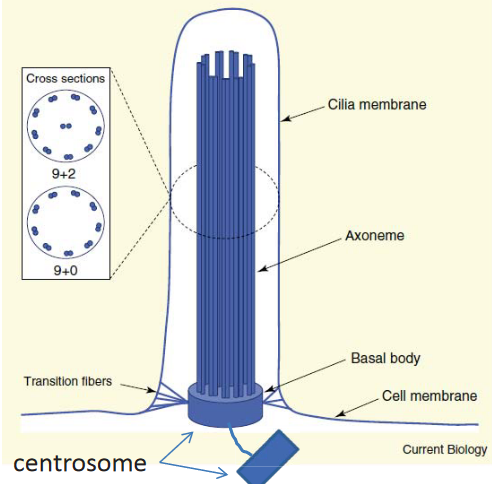
Figure 3.22: Structure of Cilia
The axoneme is a modified bundle of microtubules that extends from the basal body (i.e., the centriole).
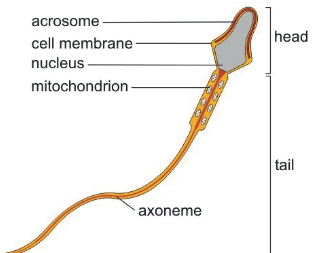
Figure 3.23: Diagram of a Sperm
Such cilia are also called flagella - these are used for movement.
Figure 3.24: Short Cilia
These are used for sensing inputs - almost all cells in the human body have cilia.
Doing so maintains distinct chemical environments for different reactions - for instance, enzymes in the golgi apparatus and the concentration of protons in the intermembrane space.