Topic 4 Protein Trafficking
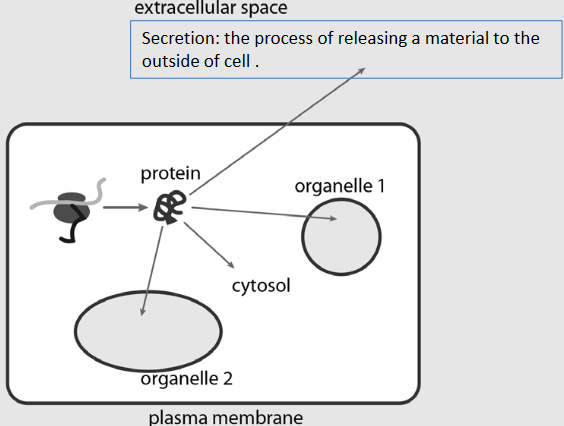
Figure 4.1: Anatomy of a Eukaryotic Cell
Among numerous membrane compartments, the burning question still remains: How does a protein know where to go and how does the protein get to its destination?
Diffusion is extremely inefficient; rather, protein trafficking is membrane mediated!
4.1 Signal Hypothesis
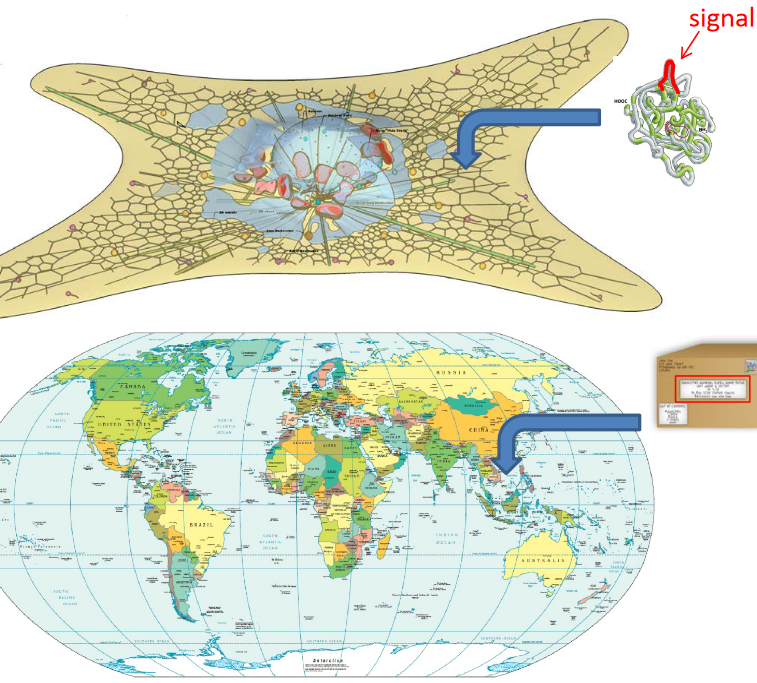
Figure 4.2: Analogy of the Signal Hypothesis
This was first proposed by Gunter Blobel; his hypothesis states that proteins have an intrinsic signal that governs its transport to its destination.
In some sense, one can think about the hypothesis as the postal address on a parcel.
4.1.1 Signal sequence
A protein that is to be sent to some secretory pathway has a signal sequence that is eventually cleaved by enzymes in the cell:

Figure 4.3: Example of a Signal Sequence
The sequence itself is attached to the N-terminus of the protein, has a short stretch of amino acids, and is long enough enough for targeting by the endoplasmic reticulum.
4.1.2 Synthesizing secretory proteins in the rough ER
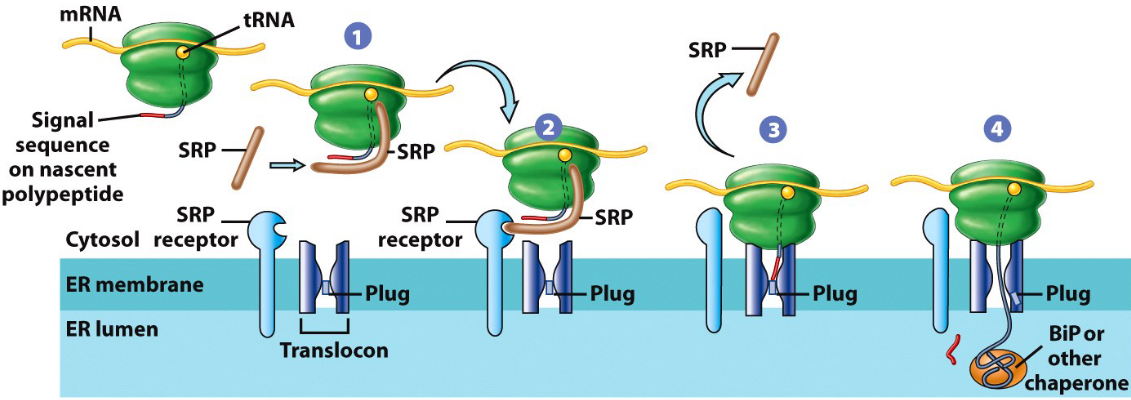
Figure 4.4: Synthesis of Secretory Proteins in the Rough Endoplasmic Reticulum
The above figure displays several steps for such a synthesis:
- The signal sequence is mediated.
- The SRP (i.e., signal recognition particle) binds to the SRP receptor.
- Co-translational translocation happens through the ER membrane.
- The signal sequence cleavage by signal peptidase in the ER lumen.
4.2 Protein Export
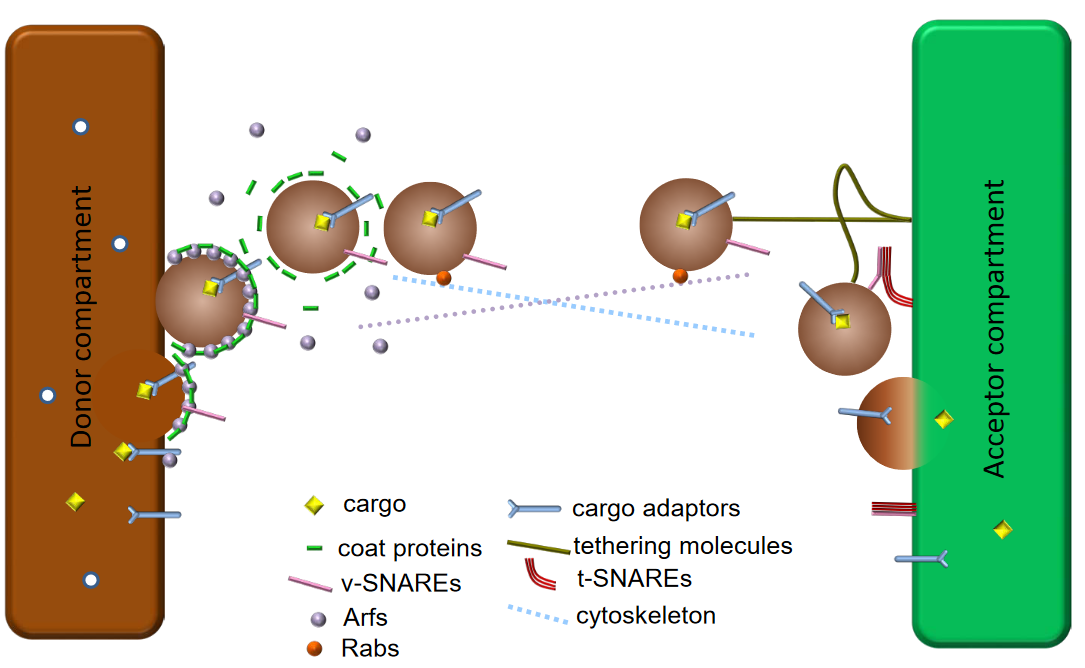
Figure 4.5: Professor Lei’s Vesicular Trafficking Model
The model suggests that vesicular trafficking is reversible - that is, anterograde and retrograde trafficking can occur:
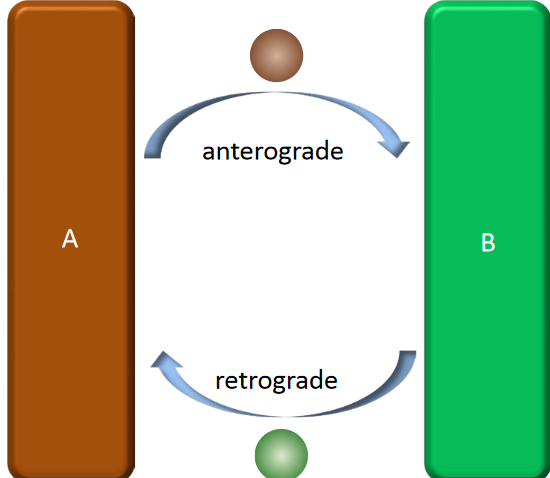
Figure 4.6: Anterograde and Retrograde Trafficking
Anterograde and retrograde trafficking refers to retrieving and recycling mechanisms.
The specificity of the above is achieved by tethering and v-t SNARE pairing. In both forms of trafficking, trafficking is reversible and also involves extensive membrane re-modelling!
4.2.1 Vesicle coats
Vesicle coats exist to mechanically bend the membrane and cargo protein selection.
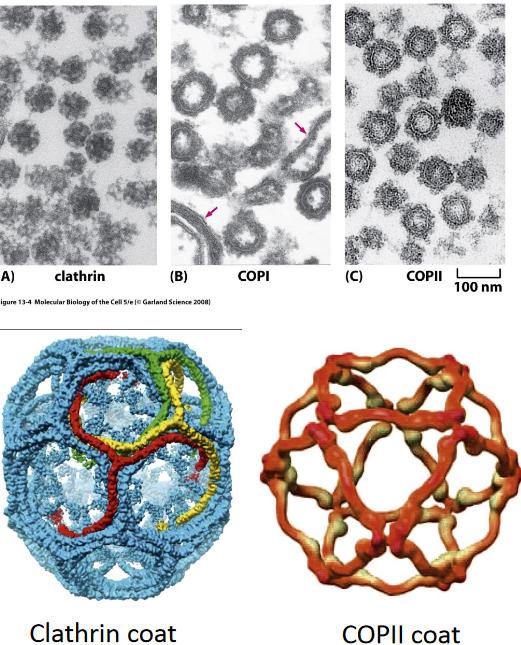
Figure 4.7: Three Types of Vesical Coats
The three types of vesicle coats examined in BS2004 are:
- COPI
- COPII
- Clathrin
All three coats are symmetrical and composed of a few types of subunits. All three coats are about 50 - 100 nm in size.
4.2.1.1 Cargo exiting COPII coated vesicles
COPII-coated vesicles are generated at the exit sites of the endoplasmic reticulum.
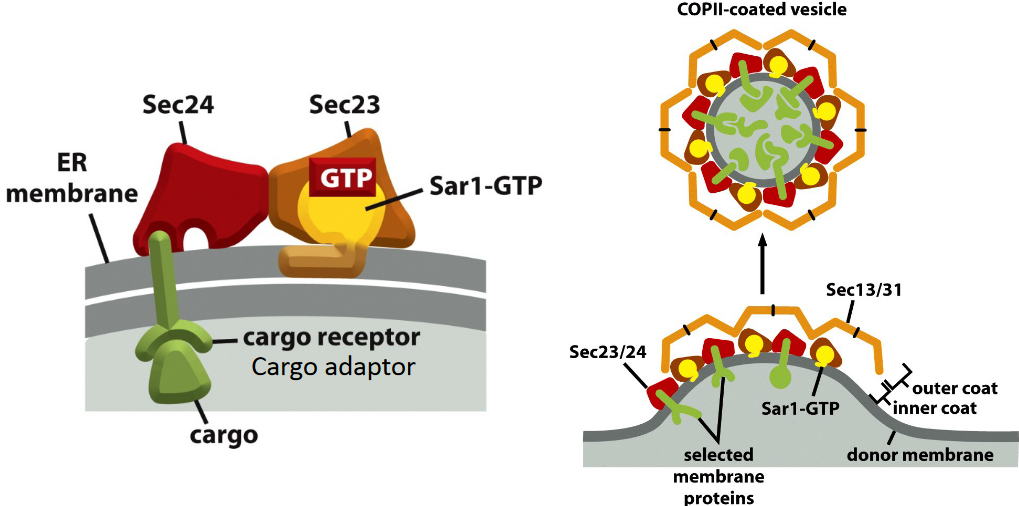
Figure 4.8: Cargo Exiting the ER in COPII-Coated Vesicles
Also important is that the Sar1 GTPase is essential for the assembly of the COPII coat.
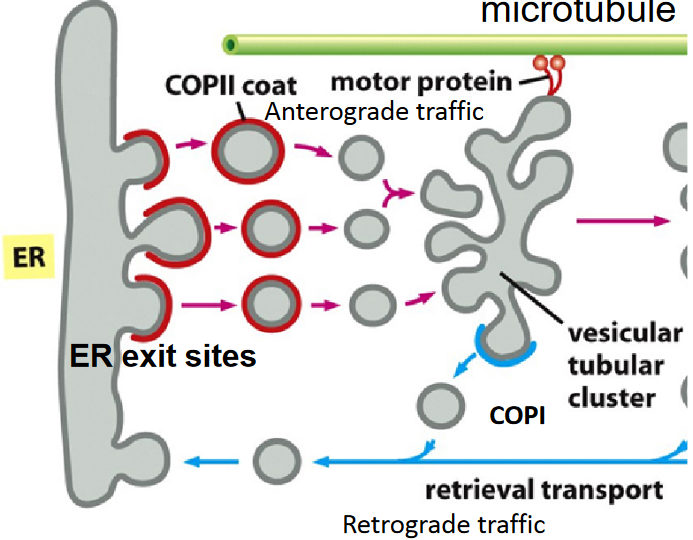
Figure 4.9: Retrograde Traffic During COPII Vesicle Export
4.2.1.2 Maturation model
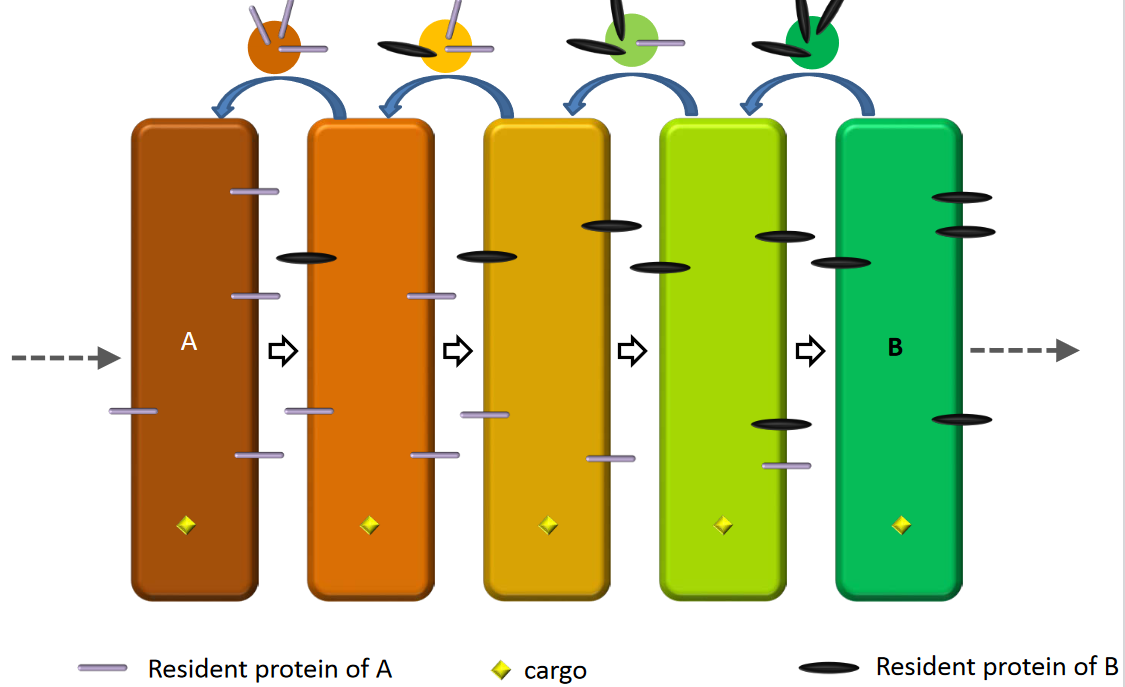
Figure 4.10: Maturation Model
The model also shows how cargo can be transported through the Golgi complex:
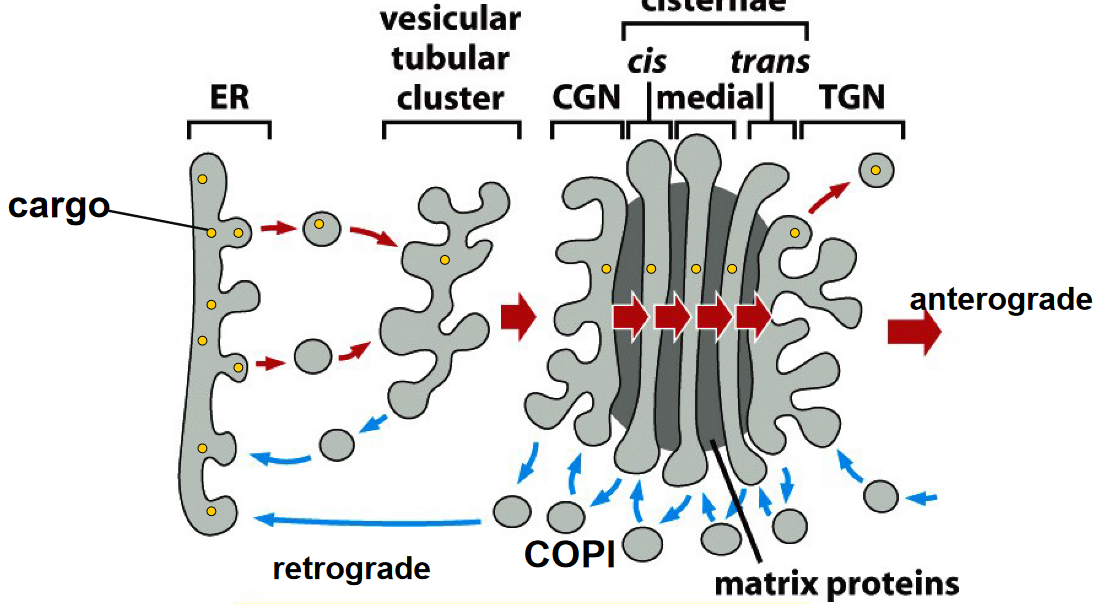
Figure 4.11: Cargo Transport via Maturation Model
4.2.1.3 Secretory pathway
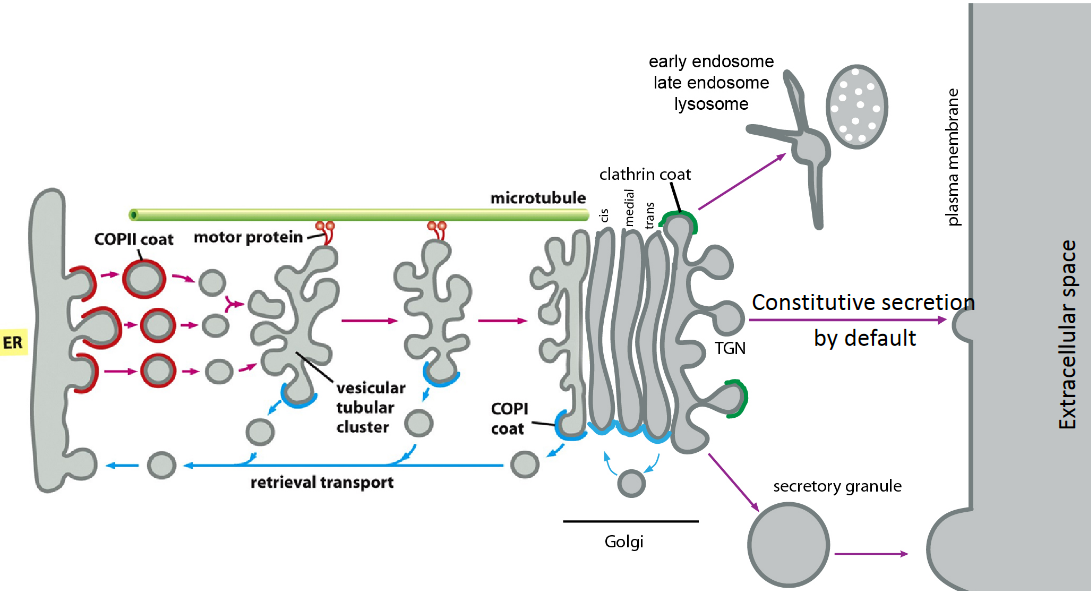
Figure 4.12: Diagram of the Secretory Pathway
The kinetics present in this pathway is very fast: about 10 minutes from the ER to the golgi.
4.2.1.4 Memrbane topology during trafficking
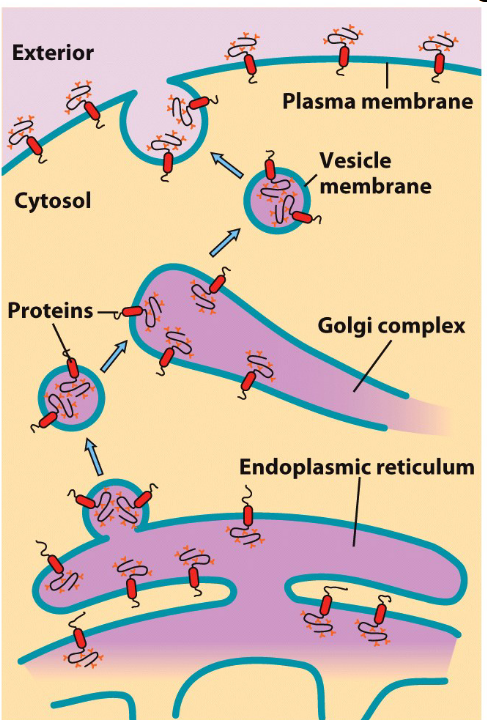
Figure 4.13: Membrane Trafficking Diagram
When membrane trafficking happens, the transmembrane proteins’ and lipids’ topologies are also conserved! This statement applies to secretory and endocytoic pathways.
The above is due to the flip-flop nature of the transmembrane proteins. Lipids are prohibited from flip-flopping.
4.2.2 Endocytosis and exocytosis
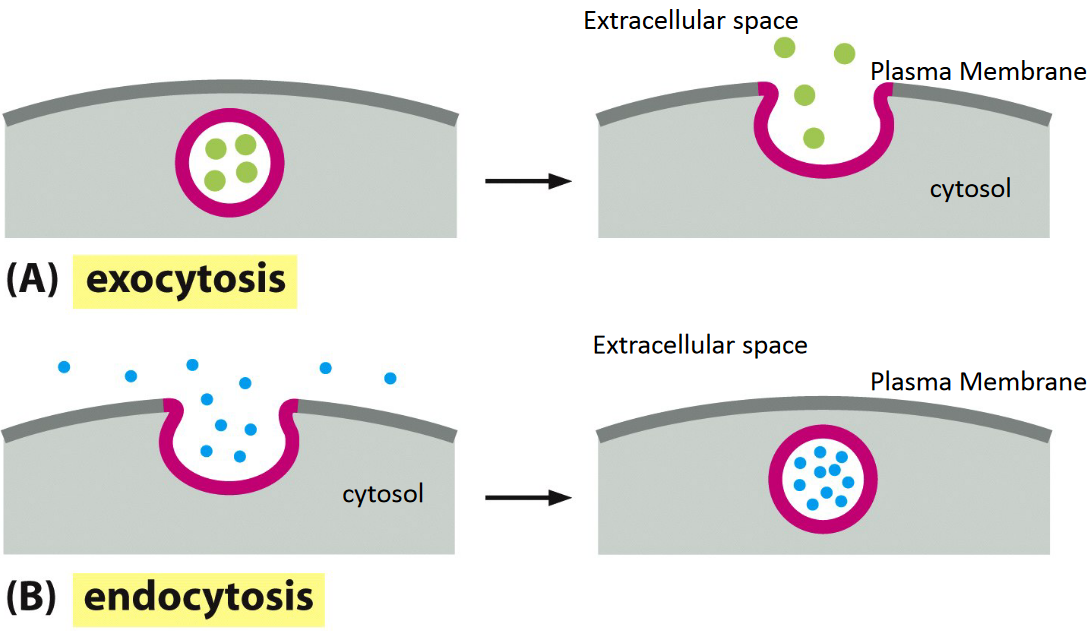
Figure 4.14: Comparison of Endocytosis and Exocytosis
4.2.2.1 Clathrin dependent endocytosis
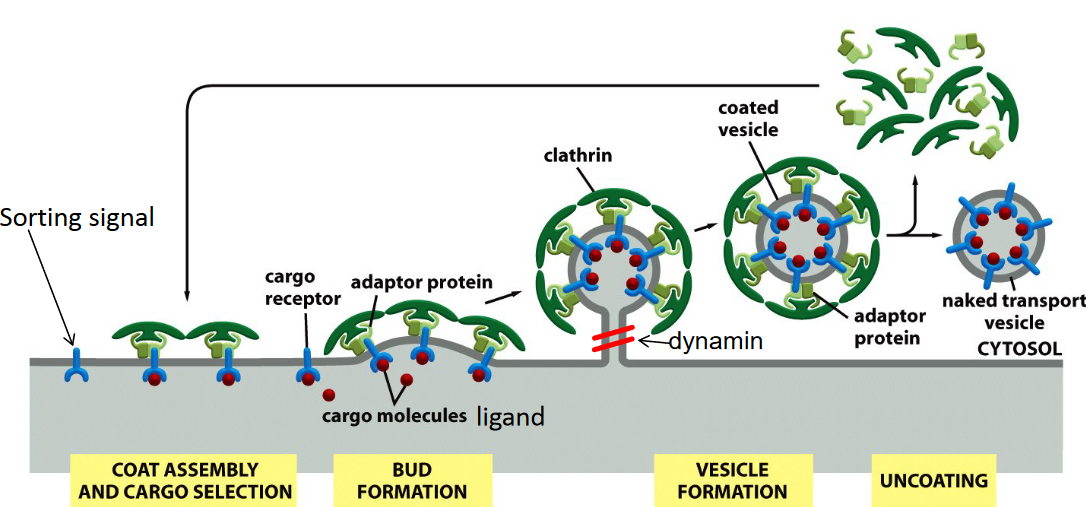
Figure 4.15: Clathrin-Dependent Endocytosis
4.2.2.2 Endocytosis of LDL/LDL receptor
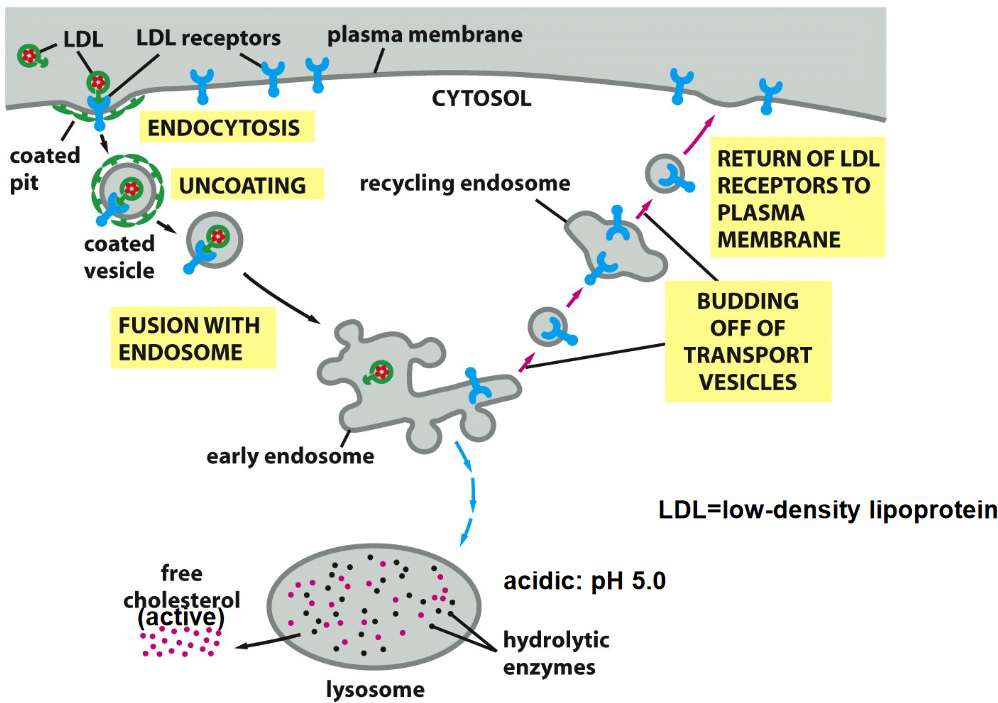
Figure 4.16: Diagram Representing the Endocytosis of the LDL/LDL Receptor
4.2.2.3 Endocytosis of EGF/EGF receptor
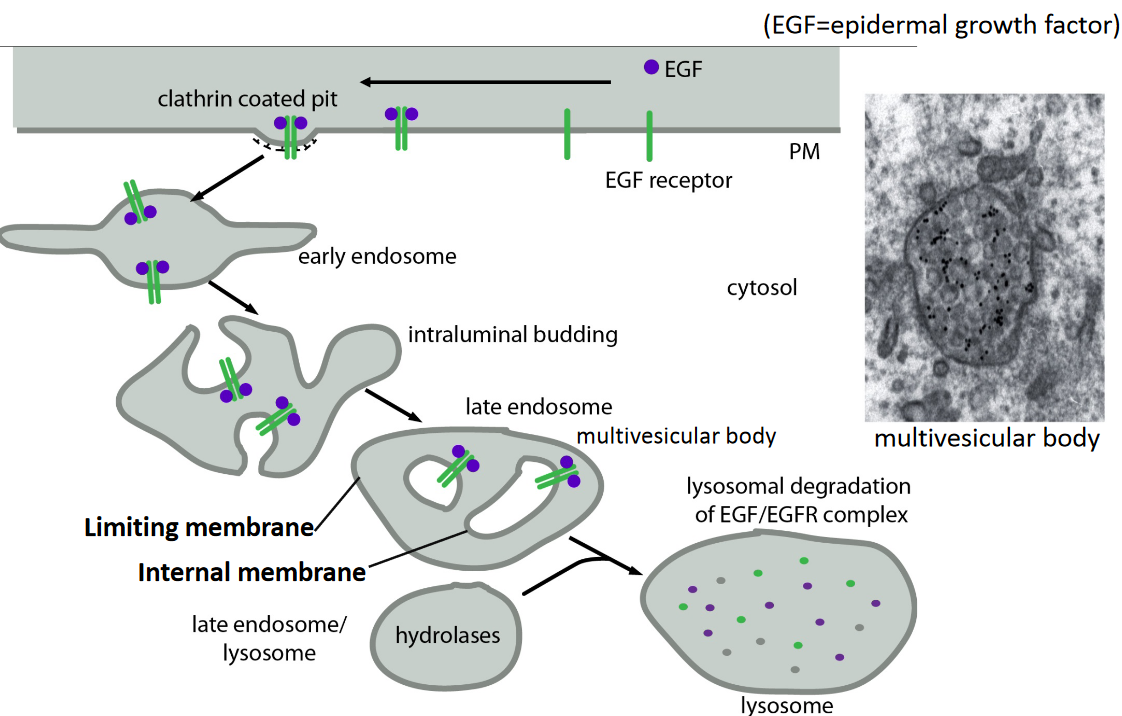
Figure 4.17: Diagram Representing the Endocytosis of the EGF/EGF Receptor
4.3 Phagocytosis and Autophagy
Phagocytosis and autophagy in cells also lead to a degradation in lysosomes
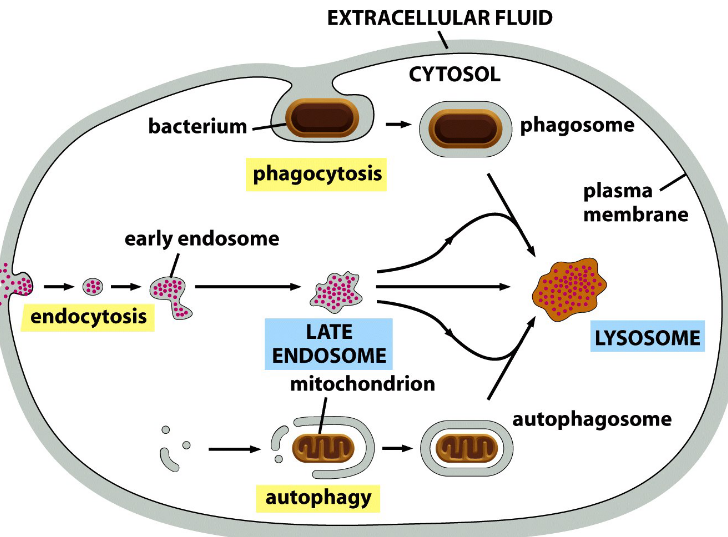
Figure 4.18: Phagocytosis and Autophagy Degrading Lysosomes
Phagocytosis happens in special cells and is a special form of endocytosis that involves the formation of a phagosome: a large endocytotic vesicle. Here, the phagosome fuses with the lysosome and the enclosed contents of the phagosome are digested!
Autophagy is used to dispose obsolete organelles in the lysosome and leads to the formation of an autophagosome. Here, the autophagosome fuses with the lysosome and the materials of the autophagosome are digested in the organelle.
4.4 Nucleocytoplasmic Trafficking
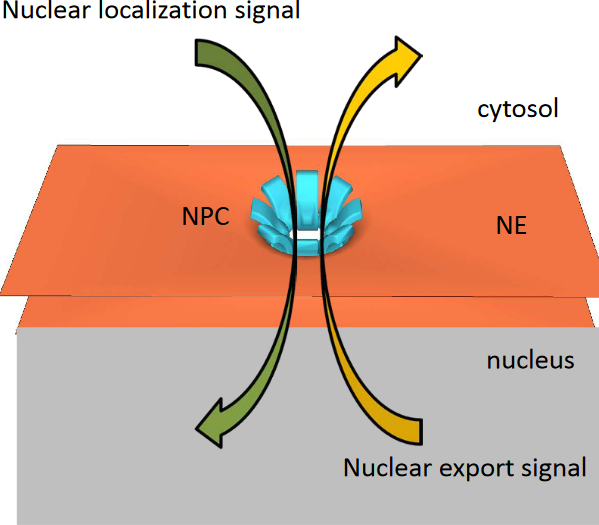
Figure 4.19: Nucleocytoplasmic Trafficking
This trafficking is signal mediated and involves two signals: a nuclear localization signal and a nuclear export signal.
In this kind of trafficking, the protein first passes through the nuclear pore complex; no vesicles are involved nor is membrane re-folding required!