Topic 8 Cell Signalling
Most cell surface receptors relay signals via small molecules and a network of intracellular signalling proteins. The number of proteins and molecules that are involved in this signalling pathway increases as the signal is propagated!
For this reason, it is no surprise that a small stimulus can lead to a huge response!
8.1 Second Messengers
Second messengers are a diverse family of compounds that are low in molecular weight: cAMP and Ca2+ ions for instance. Both these compounds transmit signals via receptor-ligand binding on the cell’s surface to induce intracellular effects - for instance, gene expression.
8.1.1 Generations of second messengers
There are two to note:
Adenylate cyclase
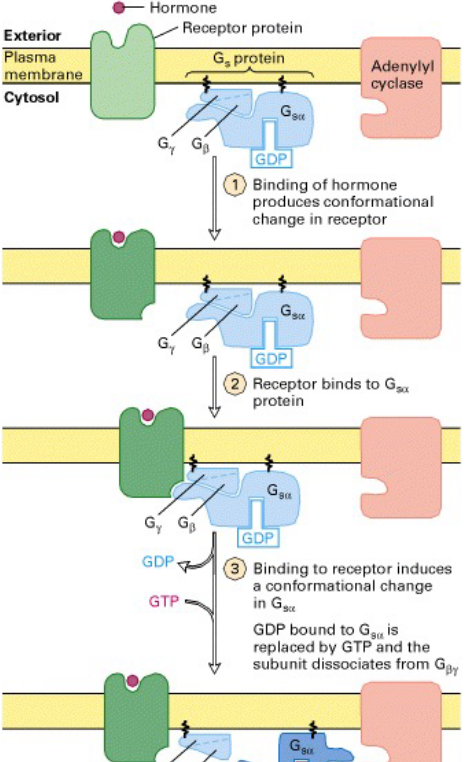
Figure 8.1: Activation of Adenylate Cyclase
This is an enzyme that catalyzes the formation of cAMP from ATP. This enzyme is also called 3’,5’-cyclic AMP synthetase.
Phospholipase C
This is an enzyme that catalyzes the formation of IP3 from PIP2.
8.1.1.1 cAMP signalling via PKA
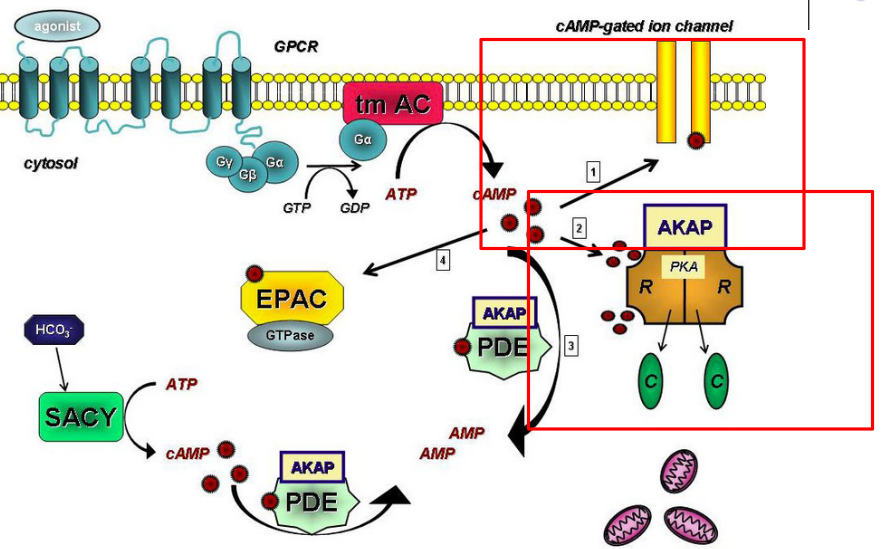
Figure 8.2: cAMP Activation via PKA
Although cAMP is capable of activating certain ion channels in the plasma membrane of most cells, it generally exhibits its effects via protein kinase a (i.e., PKA). PKA catalyzes the transfer of the \(\gamma\) phosphate group from ATP to specific threonines or serines of target proteins, hence regulating their activities.
8.1.1.1.1 Activating PKA
When PKA is inactive, it has two catalytic subunits and two regulatory subunits:
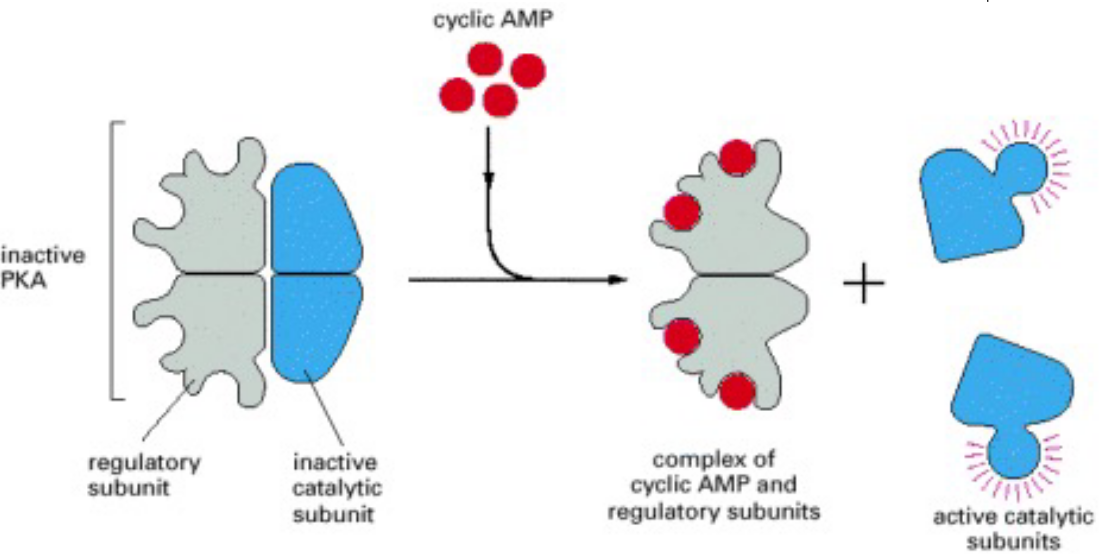
Figure 8.3: Activation of PKA
When cAMP binds to the regulatory subunits of PKA, the conformation of PKA is altered and the subunits disassociate from the complex.
8.1.1.2 Activation by phospholipase C
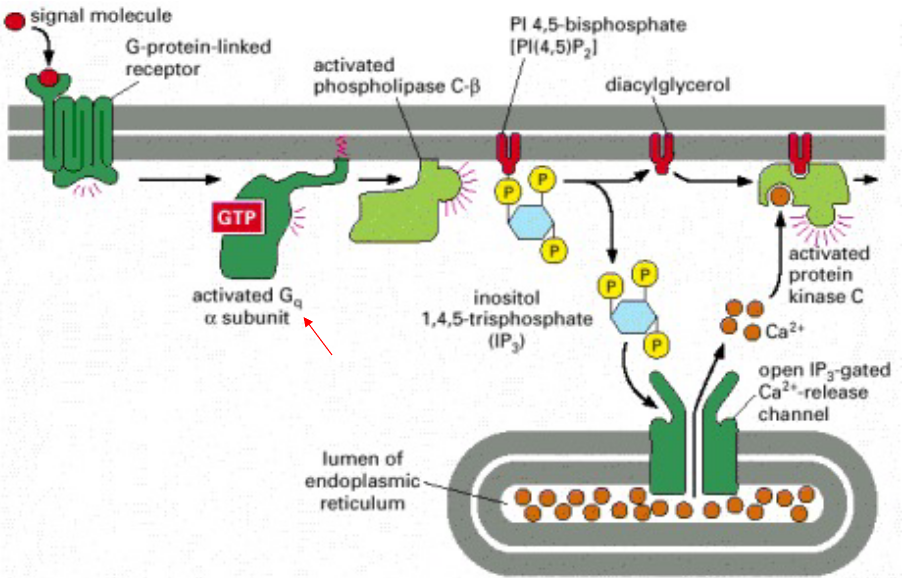
Figure 8.4: Activation by Phospholipase C
When the receptor above is stimulated, the membrane-bound enzyme phospholipase C-\(\beta\) is stimulated via a G protein. When PI(4, 5)P2 becomes hydrolyzed, two intracellular messengers are then produced.
Subsequently, IP3 diffuses through the cytosol and releases Ca2+ ions by binding to and activating IP3 gated Ca2+ ions. The large electrochemical gradient for Ca2+ allows it to escape into the cytosol.
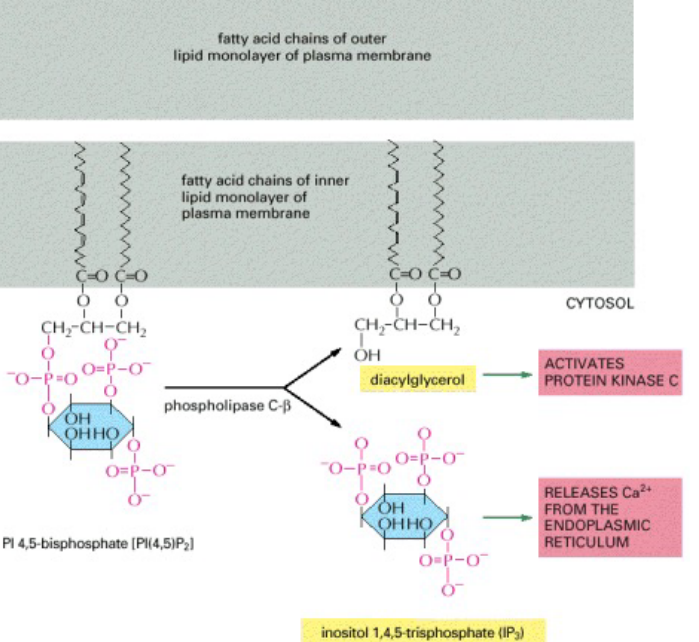
Figure 8.5: Hydrolysis of PIP2 by Phospholipase C
In the meantime, diacylglycerol remains in the plasma membrane and together with phosphatidylserine and Ca2+ ions, help to recruit the enzyme protein kinase C (i.e., PKC).
8.1.2 More on PKC
PKC is a family of protein kinases that are divided into three subfamilies:
Novel
These do not require Ca2+ ions for activation, but diacylglycerol for activation.
Conventional
These require Ca2+, a phospholipid, and diacylglycerol for activation.
Atypical
These do not depend on diacylglycerol or Ca^2+ ions for activation.
At least 15 members of the PKC family have been identified in mammals via their high sequence homology.
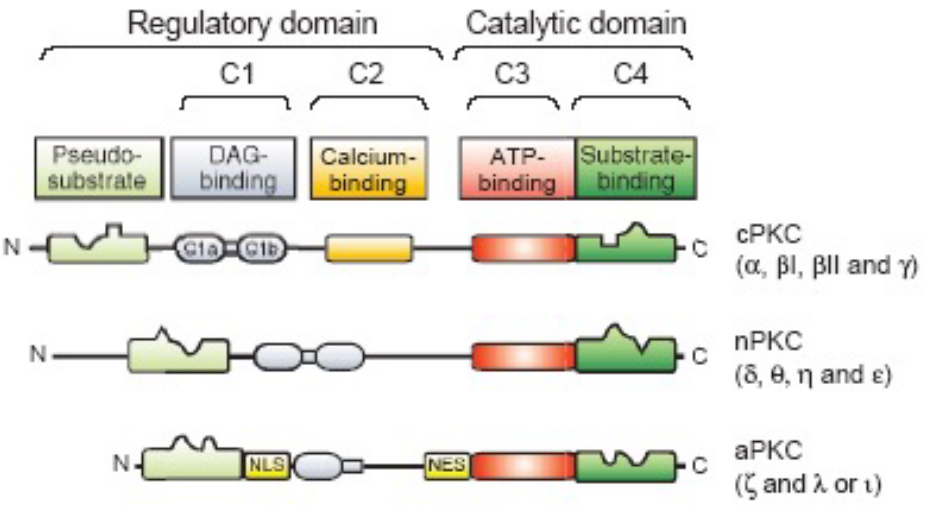
Figure 8.6: Isoforms and Regulatory Sequences of PKC
“PKC” generally refers to the protein kinase C\(\alpha\) enzyme.
8.1.2.1 PKC maturation
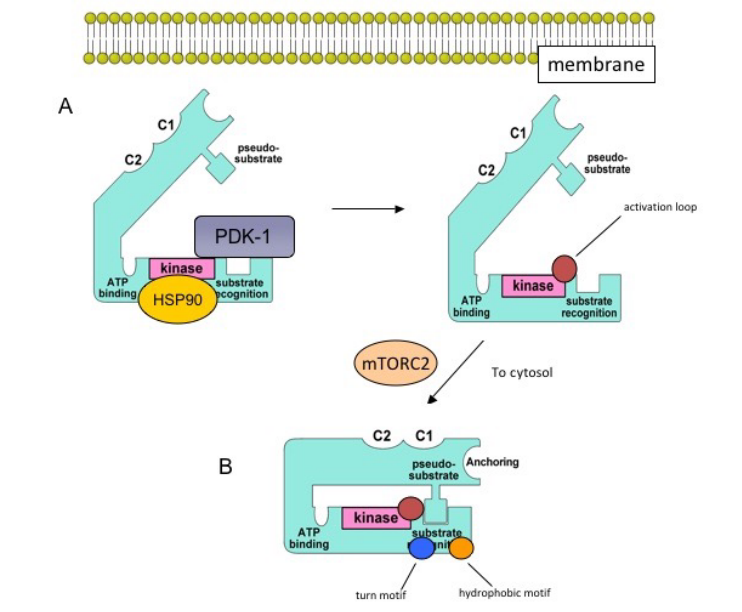
Figure 8.7: PKC Maturation Model
PKC is generally found in an inactive, open conformation that is associated with membrane fractions.
First, the HSP90 chaperone binds to the kinase domain of the inactive PKC while PDK-1 binds the carboxy terminus and phosphorylates the activation loop (shown in the circle).
The protein mTORC2 is then involved in phosphorylations of the turn motif (circled in blue) and the hydrophobic motif (in the orange circle).
The mature enzyme is then moved to the cytosol with the pseudosubstrate blocking the substrate-binding pocket.
8.1.2.2 PKC activation
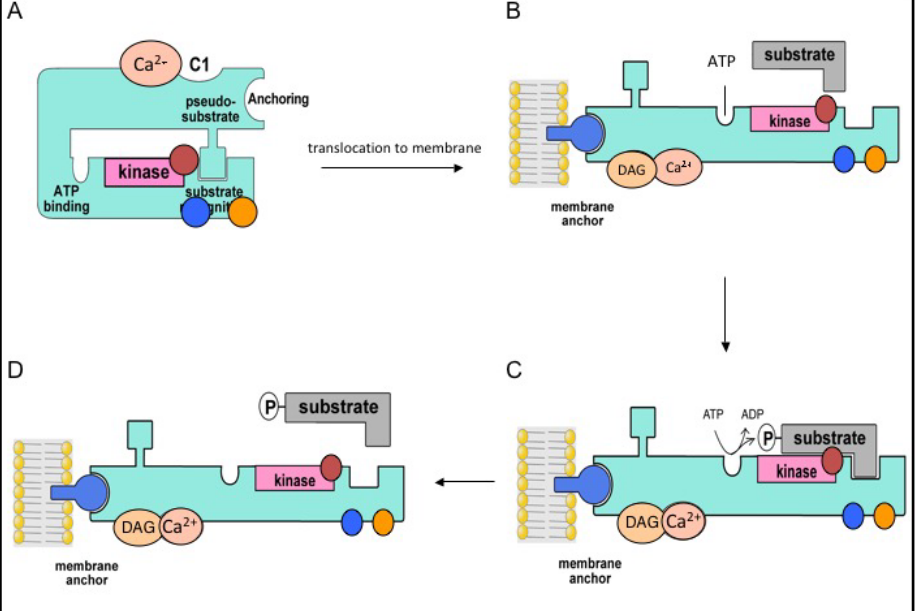
Figure 8.8: PKC Activation Model
In the inactive state “A”, the PKC enzyme is in a closed conformation.
After lipid hydrolysis, PKC is then activated via a series of steps:
- PKC binds to Ca2+ ions and translocates to the membranes.
- DAG binding occurs and promotes the activation and the opening of B - the isoenzyme.
- The activated enzyme can now bind ATP and phosphorylate various substrates.
8.1.2.3 PKC substrates
When PKC is activated, one of its target proteins can become phosphorylated - one of which is MARCKS protein. The MARCKS protein is involved in a variety of cellular processes, including but not limited to actin cytoskeleton rearrangement.
Some other targets include the MAP kinase, transcription factor inhibitor I\(\kappa\)B, and calpain.
8.1.3 CaMKII and maintaining low cytosolic Ca2+ concentrations
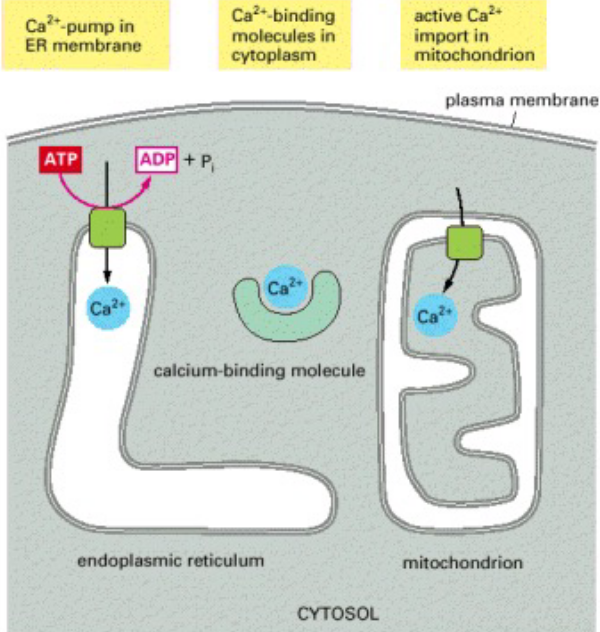
Figure 8.9: Ways of Maintaining Minimal Ca2+ Concentrations in Cells
CaMKII or Calcium almodulin kinase II transduces changes in free Ca2+ to changes in the phosphorylation status and activity of target proteins involved in neurotransmitter synthesis and release, neuronal plastiicty, and gene expression.
For this reason, CaMKII is important in the central nervous system, where it makes up about 2% of the brain’s total mass.
Calmodulin - a ubiquitous protein, is the cell’s calcium sensor and binds four calcium ions with a high affinity. The Ca2+ / calmodulin complex activates downstream targets, including CaM kinase II.
8.1.3.1 Structure of CaM-kinase II
CaMKII forms large oligomeric structures that are made up of one or more isoforms. These isoforms have anywhere between 4 to 14 isoforms in its assembly
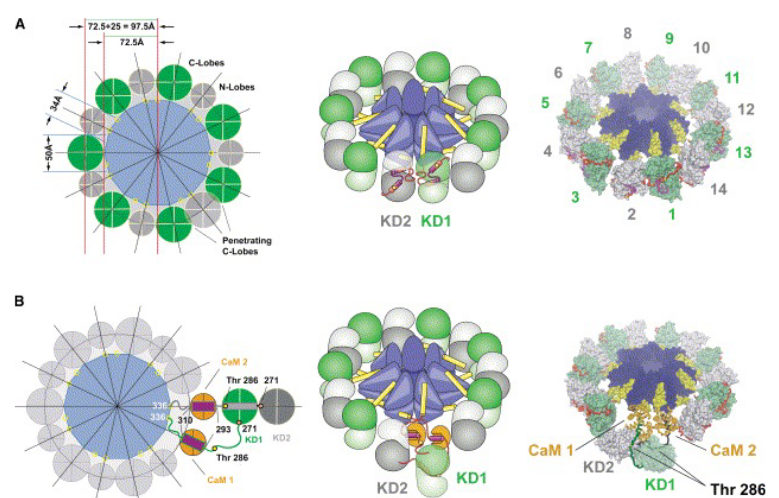
Figure 8.10: Tetrameric Isoforms of CaMKII
There are four subunits of CaMKII: \(\alpha\), \(\beta\), \(\gamma\), and \(\delta\), each of which is expressed in different concentrations in cells.
Each isoform in CaMKII also has an N-terminal serine / threonine kinase domain and a C-terminal association domain that causes oligomerization.
8.1.3.2 Activating CaMKII
When Ca2+ or calmodulin is missing, then CaMKII is inactive as a result. When Ca2+ or calmodulin binds, the protein’s conformation is altered such that the catalytic domain can phosphorylate the inhibitory domain of neighboring subunits (and other proteins in the cell).
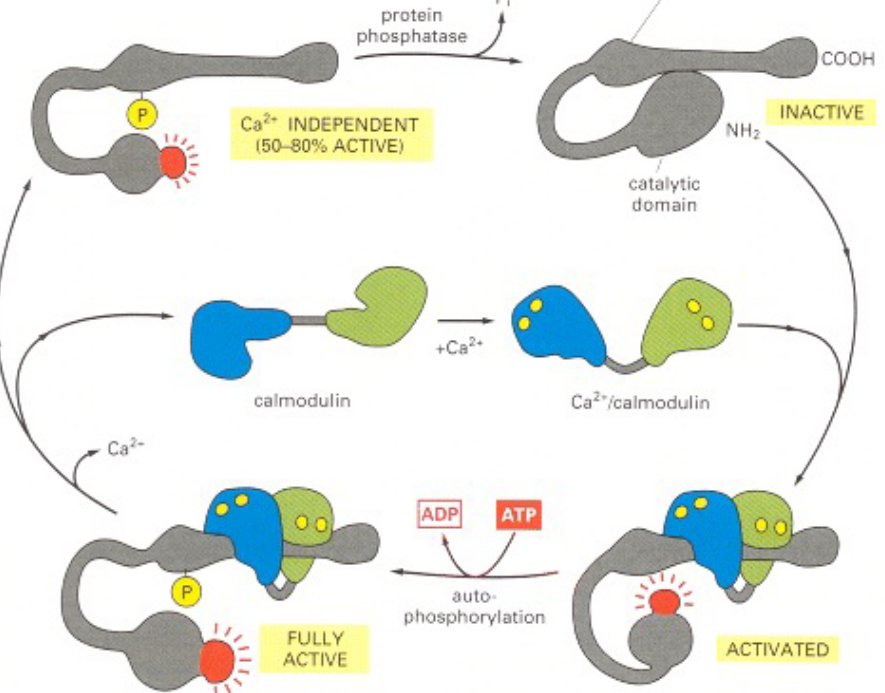
Figure 8.11: CaMKII Activation
Autophosphorylation of CaMKII can prolong the activity of CaMKII in two ways:
The Ca2+ ion or calmodulin is trapped - this way, the ions don’t disassociate from the enzyme complex until cytosolic Ca2+ ions return to base level for at least 10 seconds.
CaMKII gets turned into the Ca2+-independent form so that the kinase remains active even after Ca2+ / calmodulin disassociates from it. This continues until a protein phosophatase comes into action.
8.2 Protein Kinases
Protein kinases are a group of proteins that modify other proteins via phosphorylation. This usually results in the functional change of the target protein via changing enzyme activity, cellular location, or association with other proteins.
Up to 30% of proteins are modified by kinase activity - kinases are known to regulate the majority of cellular pathways (especially those in signal transduction). The human genome has about 500 genes that encode for protein kinases - protein kinases also constitute about 2% of all eukaryotic genes.
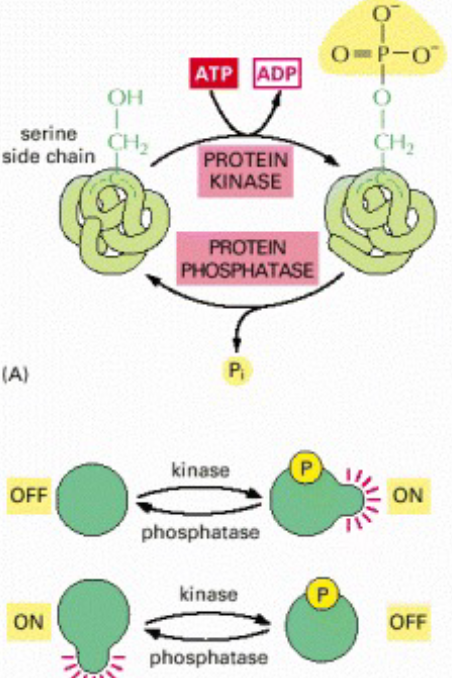
Figure 8.12: Protein Phosphorylation
Kinases do their job via removing a phosphate from ATP and attaching it to one of three amino acids that have a free hydroxyl group. Most kinases act on serine or threonine; some act on tyrosine, and a number act on all three.
8.2.1 Targeting PKC
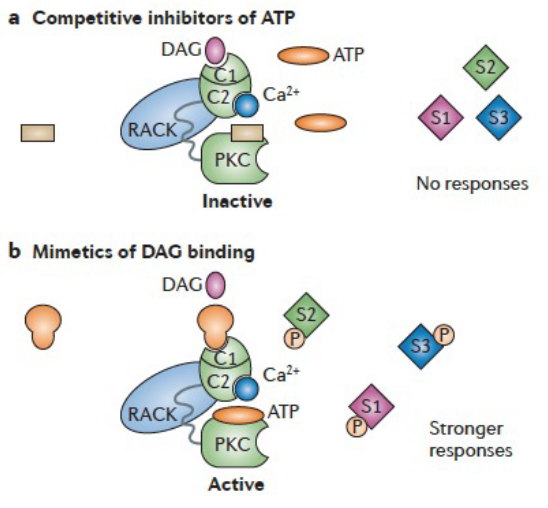
Figure 8.13: Inhibitions of PKC
Inhibitors of ATP may prevent it from binding to PKC for its phosphorylation.
PKC regulators that target the diacylglycerol binding site can also serve as activators or inhibitors.
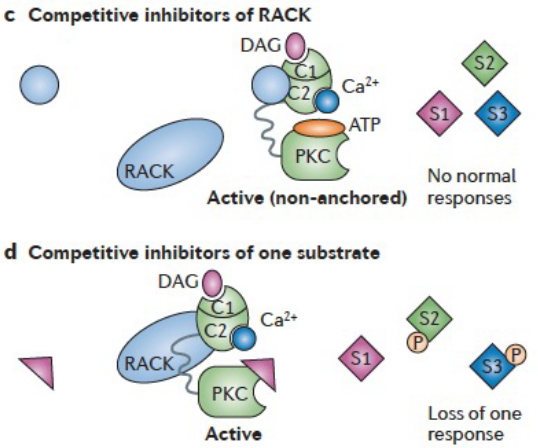
Figure 8.14: More Inhibitions of PKC
Competitive inhibitors can also stop CaMKII from anchoring to its receptor of activated C-kinase (i.e., RACK) which brings the activated isoenzyme next to its substrates - this leads to the inhibition of all physiological responses of that isoenzyme.
Otherwise, inhibitors of protein-protein interactions at a specific location or a specific substrate can also provide unique inhibitors of the phosphorylation of one substrate and not others.
8.3 Signalling Networks
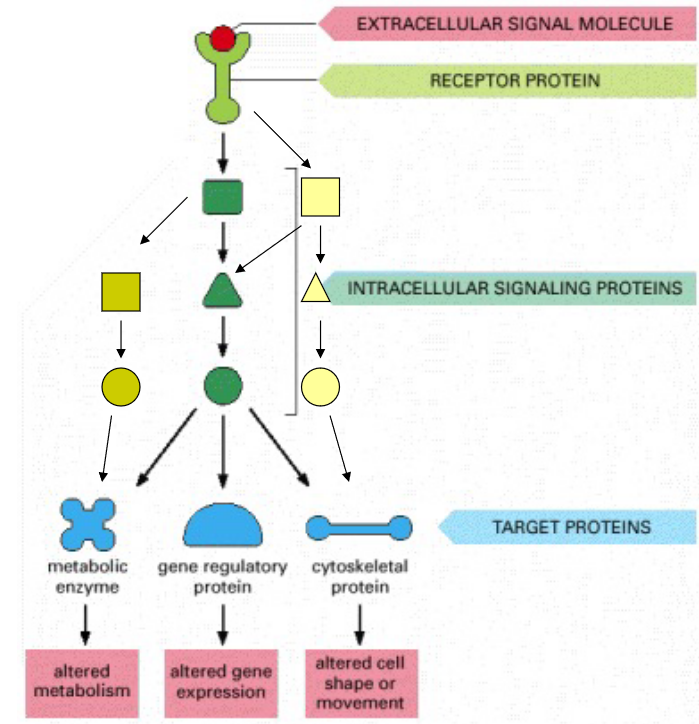
Figure 8.15: A Hypothetical Signalling Network
Signals that are received at the surface of a cell via G-protein-linked or enzyme-linked receptors are relayed into the cell interior by a combination of small and large intracellular signalling molecules.
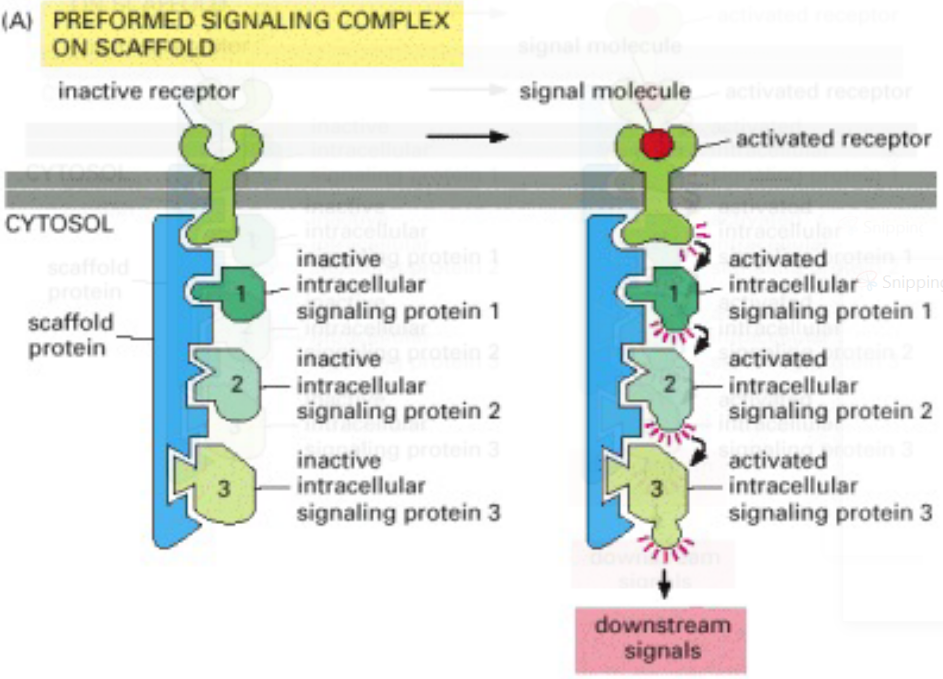
Figure 8.16: More Examples of Signalling Networks (Involving a Scaffold)
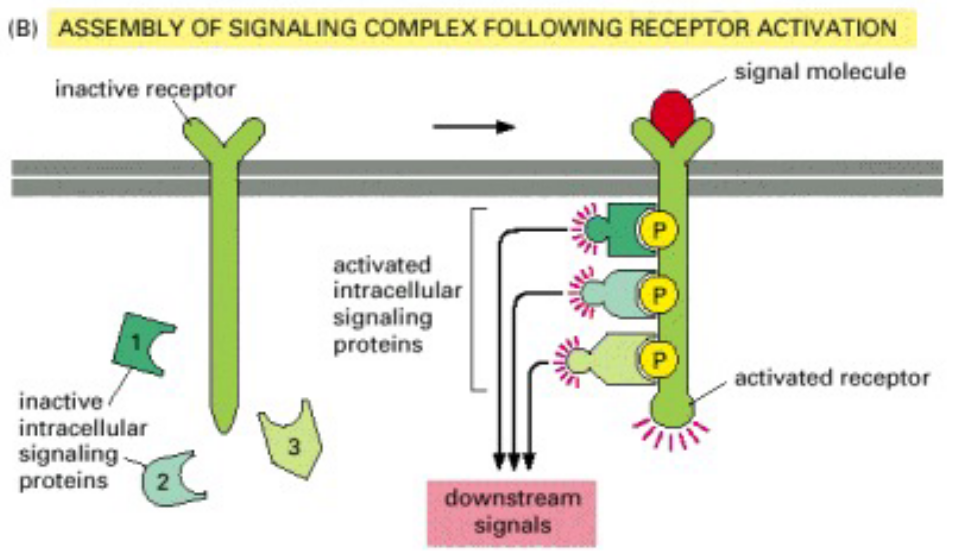
Figure 8.17: More Examples of Signalling Networks (Involving Receptor Activation)
This resulting chain of intracellular signalling events trigger target proteins - these altered proteins then modify the behavior of cells.
8.3.1 Transduction of signals via G\(_{\beta\gamma}\)
When acetylcholine binds via the muscarinic acetylcholine receptors, this triggers G protein transduction via catalyzing GTP - GDP exchange on the \(\alpha\) subunit. This released G\(_{\beta\gamma}\) subunit then opens up a K+ ion channel.
This K+ permeability increase then hyperpolarizes the membrane and reduces the frequency of heart muscle contractions.
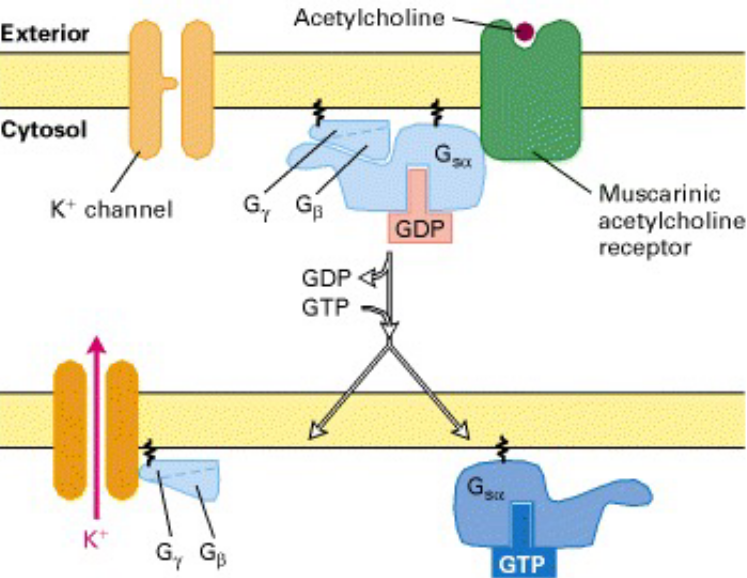
Figure 8.18: Slowing of Heart Contractions via Muscarinic Acetylcholine Receptor in the Heart
Though not shown above, when GTP is bound to G\(_{\beta\gamma}\), it becomes hydrolyzed to GDP and G\(_\alpha\)-GDP recombines with G\(_{\beta\gamma}\).
8.4 Mating in S. cerevisiae
S. cerevisiae has two main cell types: a and \(\alpha\). a cells have an \(\alpha\) factor receptor that secretes an a factor. On the contrary, \(\alpha\) cells have an a factor receptor that secretes \(\alpha\) factors.
8.4.1 Heterotrimeric G proteins in yeast mating
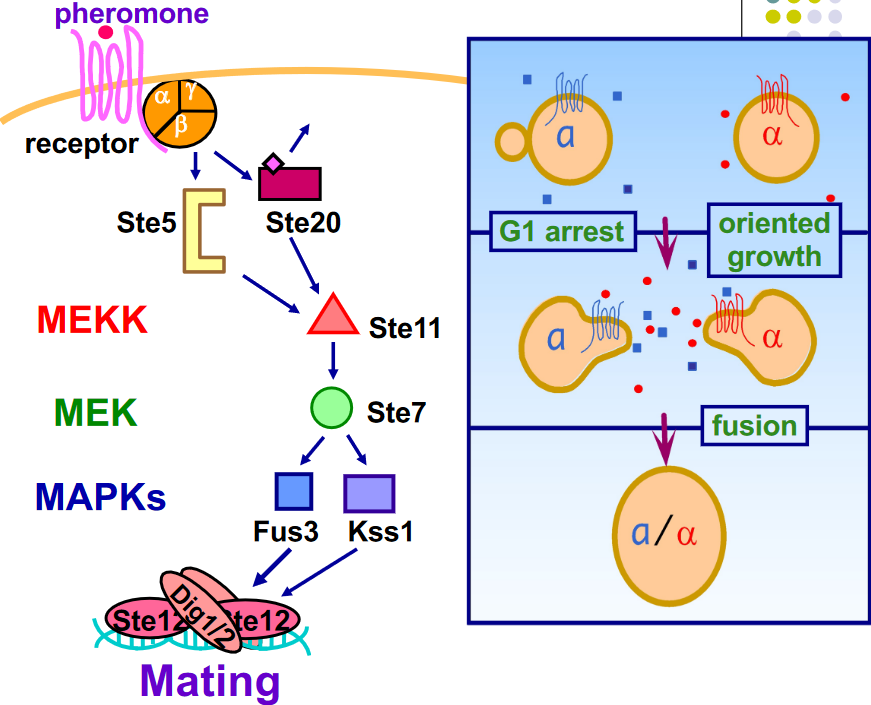
Figure 8.19: Mating Pheromone Response Pathway
Note the G-protein with the \(\alpha\), \(\beta\), and the \(\gamma\) subunits:
\(\alpha\) subunit
This is made up of GPA1 - GTP-binding protein alpha subunit of the pheromone pathway
\(\beta\) subunit
This is made up of Sterile 4 (i.e., Ste4)
\(\gamma\) subunit
This is made up of Sterile 18 (i.e., Ste18)
If either the \(\beta\) or the \(\gamma\) subunit is deleted, then the mutants are sterile and no diploids can be formed.
8.4.2 Kinases to note
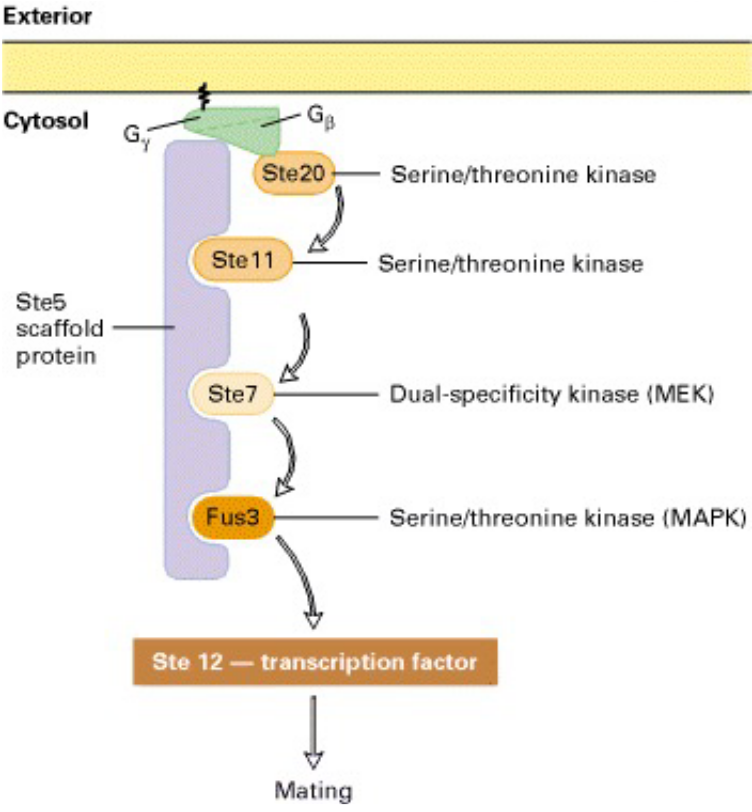
Figure 8.20: Kinase Cascade Pathway
Ste20, Ste11, Ste7, Fus3, and Kss1 are all protein kinases:
- Ste20 phosphorylates and activates Ste11
- Ste11 phosphorylates and activates Ste7
- Ste7 phosphorylates and activates Fus3 and Kss1
8.4.3 Scaffold proteins
These are proteins that reduce the extent of crosstalk and hence, maintain pathway specificity and prevent irrelevant stimuli.
Scaffold proteins have also been shown to amplify signalling.
8.5 Molecular Switches
There are two kinds:
Phosphorylation
Signals can be turned off / on by the addition or the removal of phosphate groups.
GTPase
Signals are turned on / off by the binding or the hydrolysis of GTP (to GDP) respectively
8.5.1 Desensitization to target cells
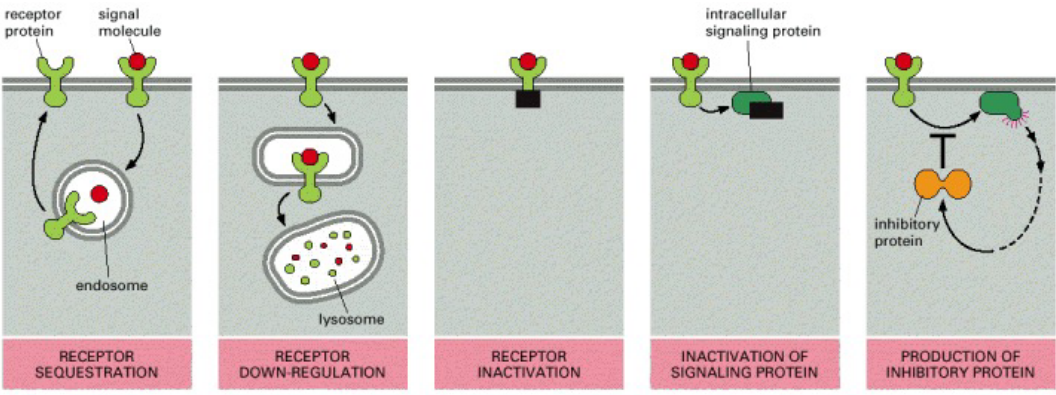
Figure 8.21: Modes of Desensitization to a Signal Molecule