Chapter 7 Other Pathways of Carbohydrate Metabolism
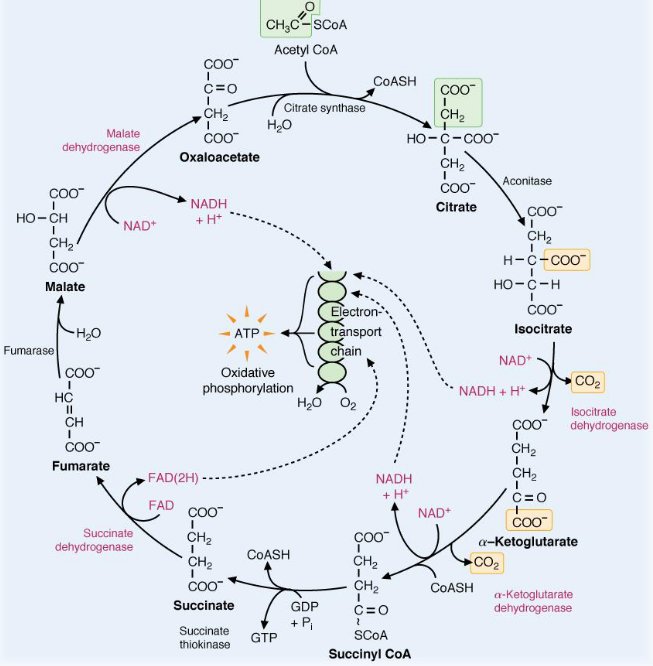
Figure 7.1: Oxidation-Reduction Enzymes in the TCA Cycle
All oxidation-reduction enzymes and coenzymes in the above TCA cycle are shown in magenta. All carbons that leave the cycle are enclosed in yellow boxes; carbons that enter the TCA cycle are enclosed in green boxes.
7.1 Glyoxylate Cycle
The glyoxylate cycle is a means of metabolizing two-carbon substrates (e.g., acetate) via eliminating CO2-producing reactions and enhancing the production of four-carbon dicarboxyl acids.
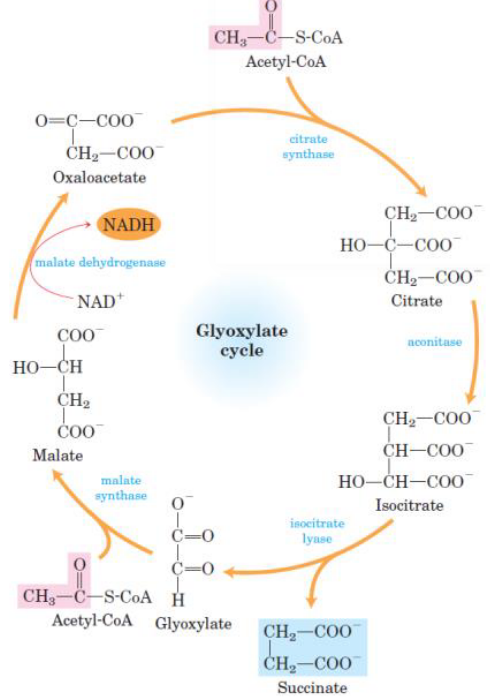
Figure 7.2: Glyoxylate Cycle
Note that the citrate synthase, the aconitase, and the malate dehydrogenase of the glyoxylate cycle are isoenzymes of the TCA cycle’s enzymes.
The reactions of the glyoxylate cycle proceed simultaneously with those in the TCA cycle. The former cycle results in the net conversion of two acetyl-CoA molecules to two succinate molecules in the glyoxysome (i.e., an organelle where the glyoxylate cycle takes place) - a product that can be used for gluconeogenesis.
In gluconeogenesis, the conversion of succinate to oxaloacetate is catalyzed by enzymes in the TCA cycle.
7.1.1 Origins of the glyoxylate cycle
Plants store lipids in their seeds - this is an energy source for plants during germination (i.e., before photosynthesis is able to take over).
Acetyl-CoA - a product from fatty-acid oxidation - is used in gluconeogenesis to produce glucose adn then sucrose and many other metabolites.
In palnt seedlings, sucrose also provides much of the chemical energy required for growth.
Plant seeds store their energy in the form of lipids rather than in carbohydrates! This is because lipids are two-fold lighter than their carbohydrate counterparts - one gram of lipids produce twice as much calories than one gram of carbohydrates.
7.2 Pentose Phosphate Pathway
7.2.1 Link between glycolysis and the pentose phosphate pathway
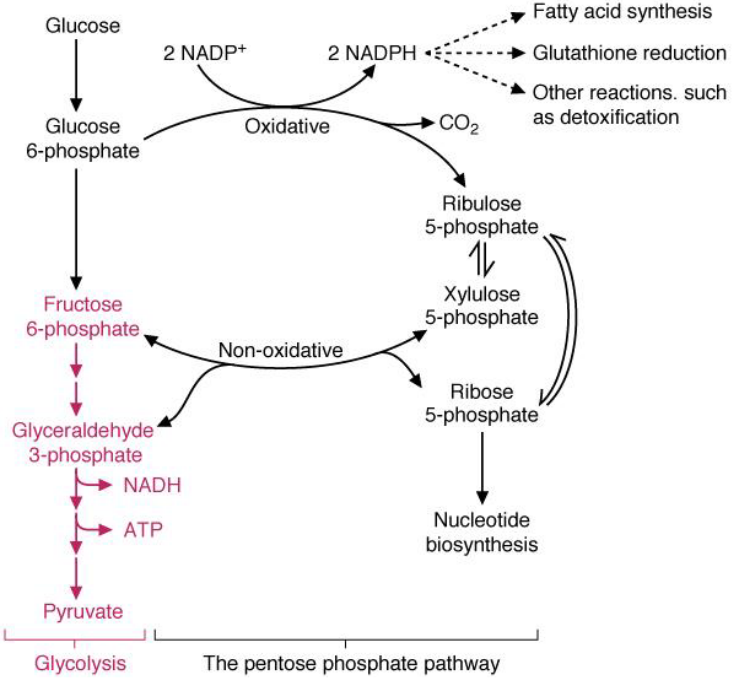
Figure 7.3: Link Between Glycolysis and Pentose Phosphate Pathway
The pentose phosphate pathway generates NADPH for reactions that require ribose-5-phosphate or reducing equivalents. The substrate glucose-6-phosphate is a substrate for both the pentose phosphate pathway and the pentose phosphate pathway.
Note that enzymes in the pentose phosphate pathway are particularly abundant in the cytoplasm of liver and adipose cells.
7.2.2 More information about the pathway
The pathway itself has two phases: an oxidative portion and a non-oxidative portion. NADPH and pentoses (especially ribose-5-phosphate) are the two most important products.
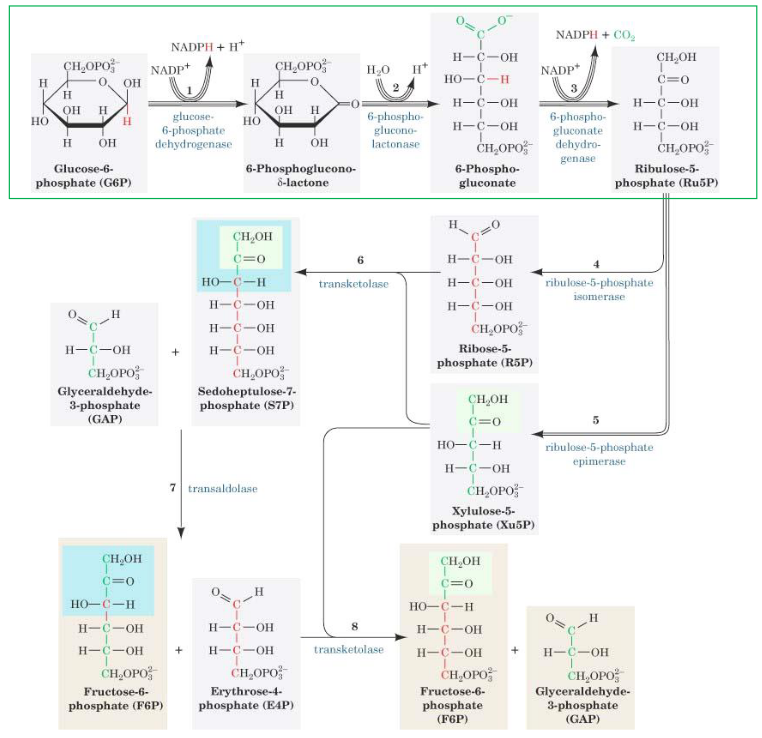
Figure 7.4: Full Pentose Phosphate Pathway
Like mentioned previously, NADH provides the reducing power for reductive pathways - for instance, fatty acid biosynthesis and the detoxification of drugs by monooxygenases.
Ribose-5-phosphate, on the contrary, is used for nucleic acid biosynthesis (which occurs at a high rate in growing and regenerating tissues and tumors). For this reason, glucose-6-phosphate dehydrogenase is the first enzyme in this pathway against oxidative damage.
7.2.3 Oxidative portion of pentose phosphate pathway
In the oxidative portion of this pathway, glucose-6-phosphate is oxidized and decarboxylated to a pentose sugar: ribulose-5-phosphate.
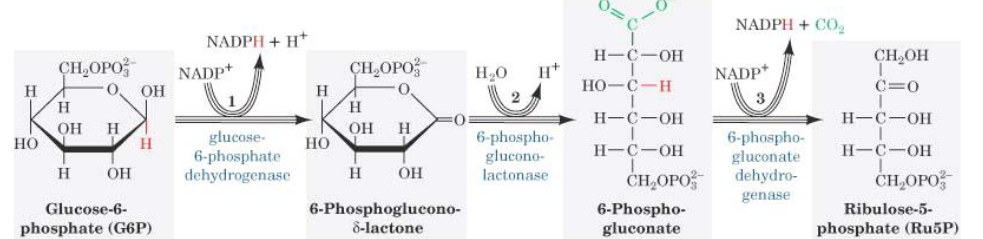
Figure 7.5: Oxidative Portion of Pentose Phosphate Pathway
Glucose-6-phosphate dehydrogenase also oxidizes the aldehyde at the first carbon and reduces NADP+ to NADPH. The gluconolactate that is formed is then rapidly hydrolyzed to 6-phosphogluconate: a sugar acid with a carboxylic acid group on the first carbon. This reaction releases CO2, reduces NADP+ to NADPH and is mechanistically similar to the reaction catalyzed by isocitrate dehydrogenase in the TCA cycle.
Note that NADPH can also be generated from several reactions in the liver and other tissues. Aditionally, NADPH cannot be fully oxidized by the electron transport chain - the ratio of NADPH and NADP+ in cells is also more than one.
7.2.4 Conversion of ribose-5-phosphate
The nonoxidative portion of the pentose phosphate pathway is comprised of a series of rearrangement and transfer reactions that convert ribulose-5-phosphate to ribose-5-phsophate to xylulose-5-phosphate. The enzymes epimerase, isomerase, transketolase, and transaldolase are involved.
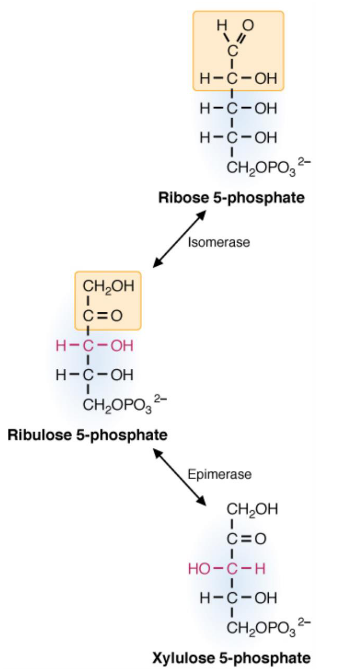
Figure 7.6: Enzyme Actions in Ribose-5-Phosphate Conversion
7.2.5 Transketolase reaction
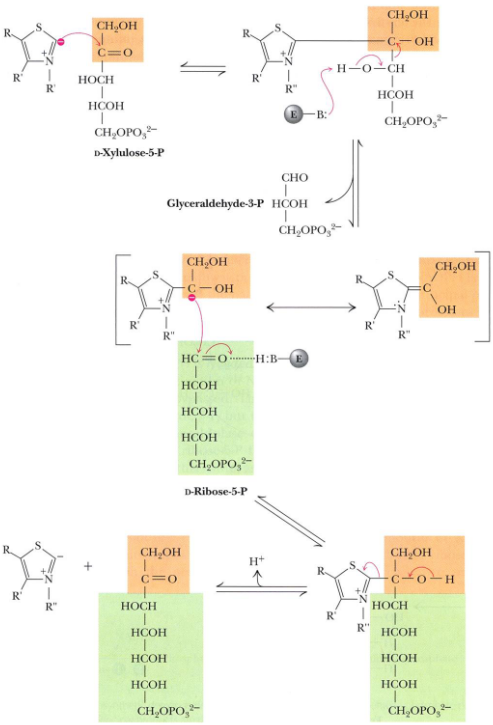
Figure 7.7: Transketolase Reaction
Transketolase in steps 6 and 8 of the pentose phosphate pathway by catalyzing the transfer of two-carbon units.
In these reactions, the donor molecule is a ketose (D-Xylulose-5-phosphate) and the recipient molecule is an aldose (Ribose-5-phosphate or Erythrose-4-phosphate).
Transketolase is also TPP-dependent - the mechanism for this enzyme involves the abstraction of an acidic thiazole proton, an attack by the carbanion at the carbonyl carbon of the ketose phosphate substrate, the expulsion of glyceraldehyde-3-phosphate, and the transfer of the two-carbon unit.
7.2.6 Transaldolase reaction
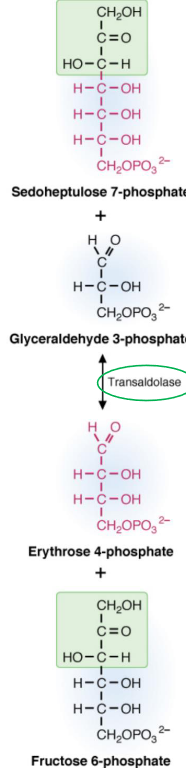
Figure 7.8: Transaldolase Reaction
Transaldolase transfers a three-carbon keto fragment from sedoheptulose-7-phosphate to glyceraldehyde-3-phosphate to form erythrose-4-phosphate to form erythrose-4-phosphate and fructose-6-phosphate. The enzymes also use an active amino group from the side chain of lysine to catalyze the reaction.
The aldol cleavage occurs between two hydroxyl carbons adjacent to the keto group. This reaction is also similar to the aldolase reaction in glycolysis.
7.2.7 Balanced reactions in the pentose phosphate pathway
The pathway ends up forming two moles of fructose-6-phosphate and one mole of glyceraldehyde-3-phosphate.
Because the pentose phosphate pathway ends and begins with glucose-6-phosphate (and feeds back into glycolysis), this is also sometimes called the hexose monophosphate pathway (HMP).
Note that the entry of glucose-6-phosphate is controlled by intracellular concentrations of NADPH - this is because NADPH is a strong product inhibitor of glucose-6-phosphate dehydrogenase.
In the liver, fatty acid synthesis from glucose is a major route of NADPH reoxidation. Synthesizing glucose-6-phosphate dehydrogenase is also induced by the increased insulin to glucagon ratio after a high-carbohydrate meal.
7.2.8 Regeneration of NADPH in the pentose phosphate pathway
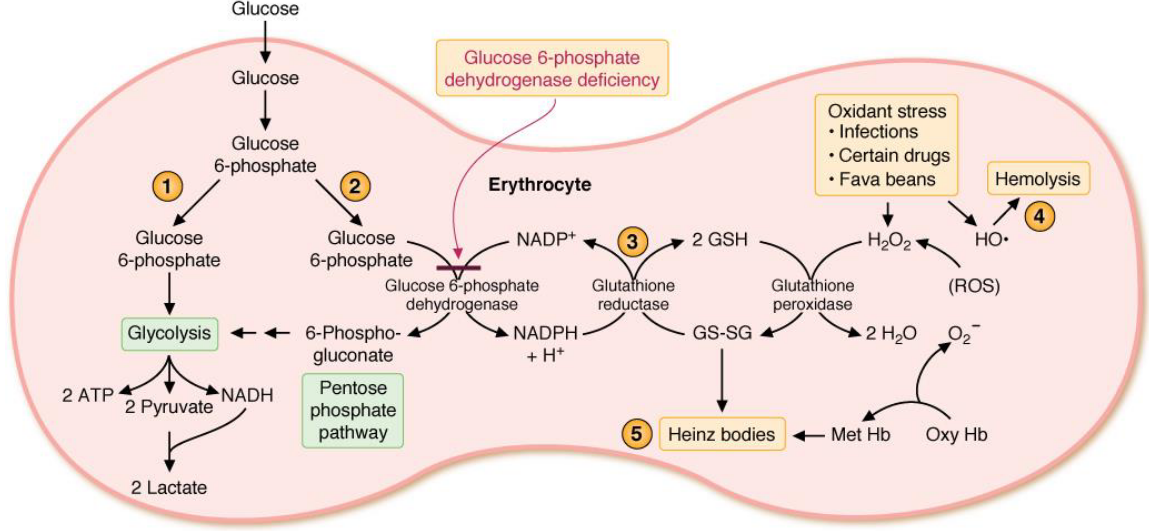
Figure 7.9: Pentose Phosphate Pathway in the Glyoxysome
NADPH - like mentioned previously - provides reducing equivalents for biosynthetic reactions and for oxidation-reduction reactions that are involved in the protection against reactive *oxygen species (i.e., ROS**).
The requirement to maintain levels of glucathione (GSH) accounts for the universal distribution of the pentose phosphate pathway throughout different types of cells.
NADPH is used as a cofactor for many reactions: fatty acid synthesis, cholesterol synthesis, and fatty acid chain elongation.
The highest concentrations of glucose-6-phosphate dehydrogenase are found in phagocytic cells where NADPH oxidase utilizes NADPH to form superoxide from molecular oxygen. The superoxide then generates H2O2 which in turn kills microorganisms that have been uptaken by these phagocytic cells.
7.3 Fructose Conversion into C3 Molecules
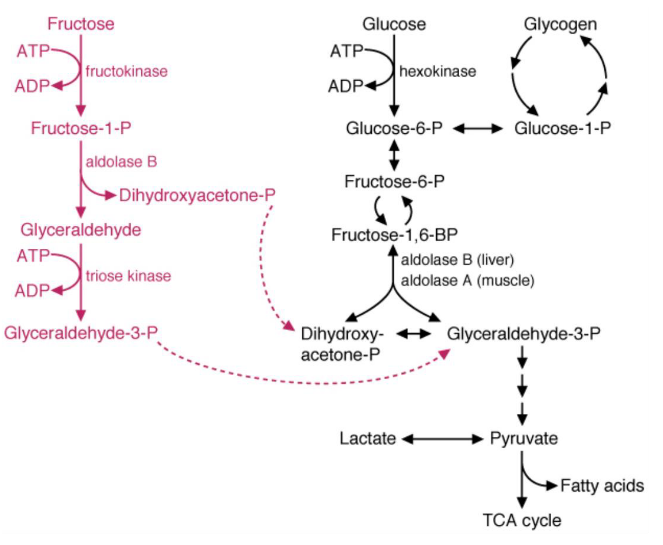
Figure 7.10: Fructose Conversion
In the first step of this conversion pathway, fructokinase phosphorylates fructose at the first carbon - the enzyme itself has a high Vmax and phosphorylates fructose as it comes into the cell.
The fructose-1-phosphate formed is then cleaved by aldolase B to dihydroxyacetone phosphate and glyceraldehyde (which can proceed to glycolysis).
The glyceraldehyde is then phosphorylated to glyceralde-3-phosphate by triose kinase.
Note that the above products can also be converted into glucose via gluconeogenesis.
7.3.1 Polyol pathway
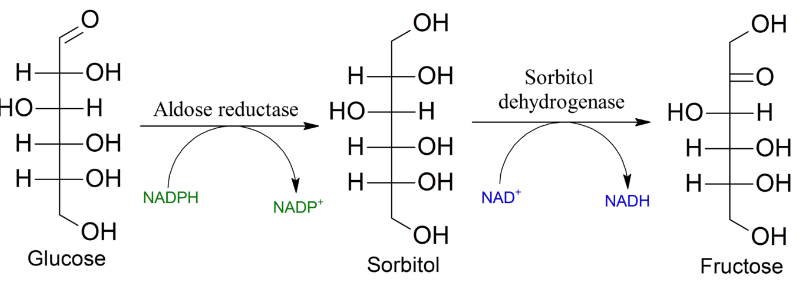
Figure 7.11: Polyol Pathway
Fructose can also be created via the polyol pathway (from glucose molecules). However, in the lens of the eyes, the polyol pathway can result in the development of cataracts.
If a large amount of glucose is present in the person’s body, hexokinase becomes saturated and the glucose enters the polyol pathway (during which aldose reductase reduces it to a sorbitol).
The above reaction also oxidizes NADPH to NADP+, and sorbitol dehydrogenase oxidizes sorbitol to fructose.
While hexokinase is able to phosphorylate fructose to fructose-6-phosphate, in patients with diabetes, the reaction’s mass balance favors the production of sorbitol and hence, increased osmotic pressure in the lens.
7.4 Gluconeogenesis
The brain, the central nervous system, the testes, the renal medulla, and embryonic tissues all require glucose as their main fuel source.
However, the human brain alone requires 120 grams of glucose - the glucose content in the body’s fluids is about 20 grams, and the total glycogen storage at any one given moment is about 190 grams. The steady-state level of glucose is about 5 mM - hence, glucose must be recycled.
The above is even more important when the individual is fasting or starving - gluconeogenesis provides about 60% of total glucose on the first day of fasting.
7.4.1 Gluconeogenesis in the liver
In humans, the major precursors for gluconeogenesis are lactate, glycerol, and amino acids (in particular, alanine). Apart from three key sequences, the reactions that take place in gluconeogenesis are essentially the reverse of the reactions that take place in glycolysis.
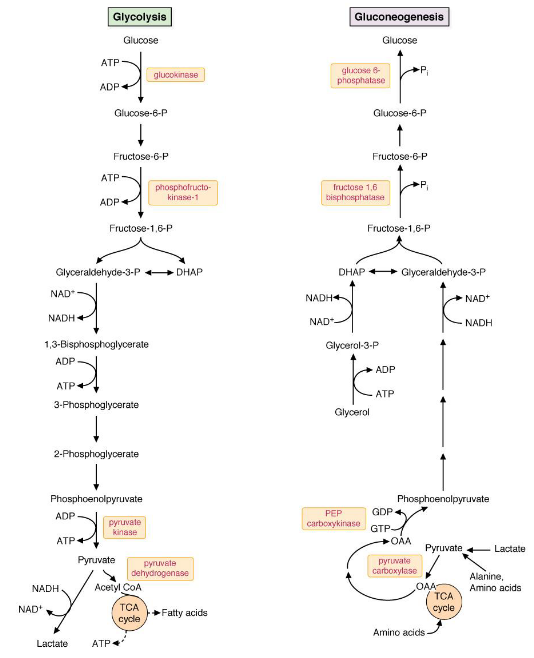
Figure 7.12: Side-by-side Comparison of Gluconeogenesis and Glycolysis
The three sequences mentioned previously are the conversion of:
- Pyruvate to phosphoenolpyruvate (i.e., PEP)
- Fructose-1,6-biphosphate to fructose-6-phosphate
- Glucose-6-phosphate to glucose.
7.4.1.1 Metabolizing gluconeogenic precursors
Lactate can be converted into pyruvate via lactate dehydrogenase (see chapter 2 of this website).
Alanine aminotransferase converts alanine to pyruvate:
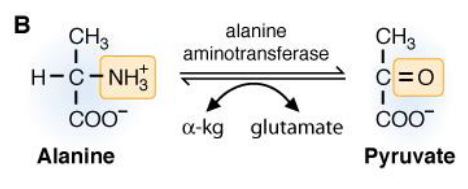
Figure 7.13: Alanine to Pyruvate
The ammonium group on the alanine molecule gets transferred over to \(\alpha\)-ketoglutarate to form glutamate. The coenzyme PLP accepts and donates the amino group.
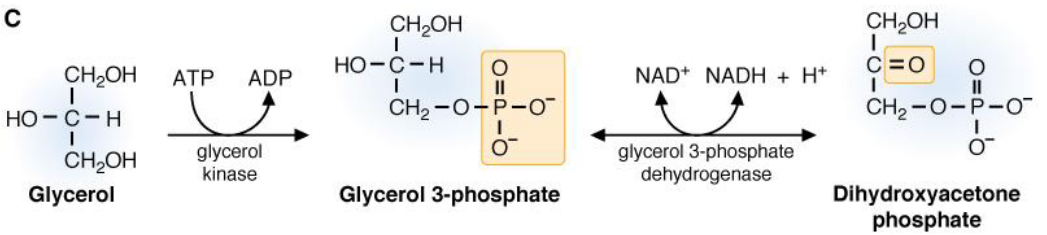
Figure 7.14: Glycerol to Dihydroxyacetonephosphate
Glycerol can also be converted over to dihydroyacetone phosphate.
7.4.1.1.1 Converting pyruvate to oxalatoacetate
This reaction is catalyzed by pyruvate carboxylase:
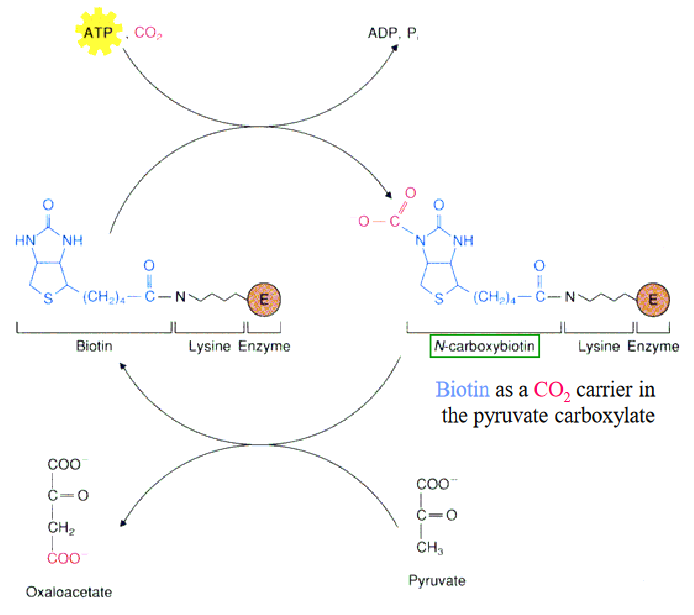
Figure 7.15: Pyruvate Carboxylase Reaction
Pyruvate carboxylase operates via a two-step mechanism and starts with the ATP-dependent carboxylation of the biotin cofactor to give N-carboxybiotin. This activated cofactor then transfers a carboxyl group directly to pyruvate.
Note that this reaction is compartmentalized - pyruvate is converted over to oxalatoacetate in the mitochondria. However, because oxalatoacetate cannot be transported across the mitochondrial membrane, it must first be reduced to malate, transported to the cytosol, and then oxidized back to oxalatoacetate before gluconeogenesis can continue.
7.4.1.1.2 Converting pyruvate to phosphoenolpyruvate
Pyruvate is carboxylated by pyruvate carboxylase:
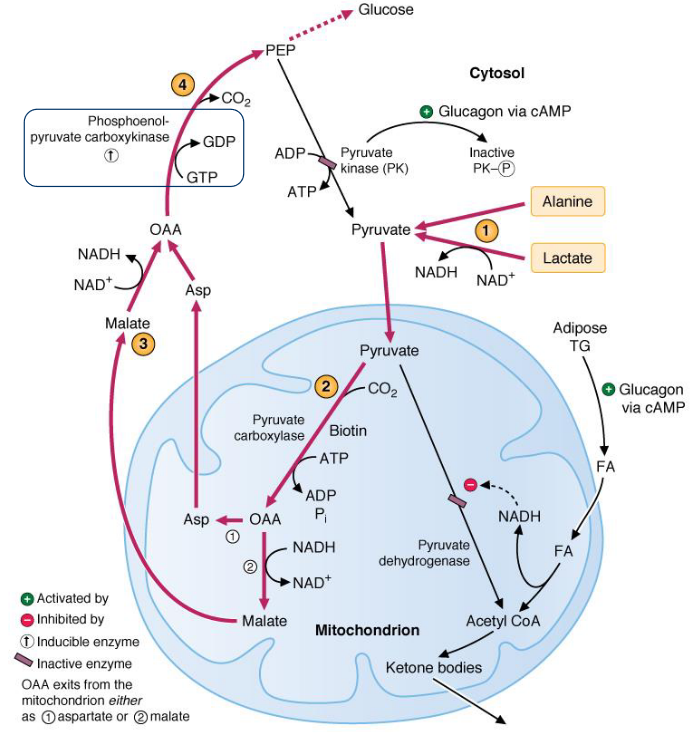
Figure 7.16: Pyruvate Carboxylase Action
This enzyme requries a biotin cofactor (for the refilling reaction of the TCA cycle). Oxalatoacetate is also replenished in this reaction and is used for the synthesis of glucose.
7.5 Miscellaneous Information
7.5.1 Fructose-1,6-biphosphate reaction
The hydrolysis of fructose-1,6-biphosphate to fructose-6-phosphate is exergonic under standard reactions.
Revisit chapter 3 for more information!
7.5.2 Glucose-6-phosphatase
This reaction requires a phosphohistidine intermediate.
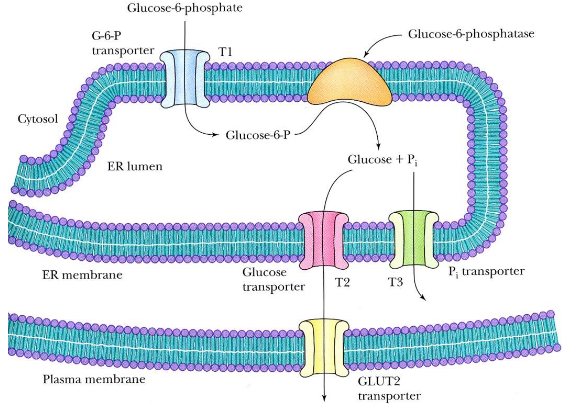
Figure 7.17: Gluconeogenesis in the Liver
This enzyme is present in the endoplasmic reticulum membrane of liver and kidney cells, but absent in muscle and brain cells. The T1, T2, and T3 transport proteins are also involved.
7.5.3 Net reaction of gluconeogenesis
Gluconeogenesis produces the following:
\[\begin{equation} 2 pyruvate + 2 NADH + 2H^+ + 4 ATP + 2 GTP + 6H_2O \rightarrow glucose + 2 NAD^+ + 4 ADP + 2 GDP + 6 P_i \end{equation}\]
The free energy change for this conversion is -37.7 kJ / mol. The consumption of six nucleoside phosphate triphosphates drives this process forward.
7.5.4 Sources of blood glucose
Right after a meal, dietary carbohydrates act as a major source of blood glucose. However, as blood glucose levels return to the fasting range within two hours after a meal, glycogenolysis starts supplying glucose into the blood:
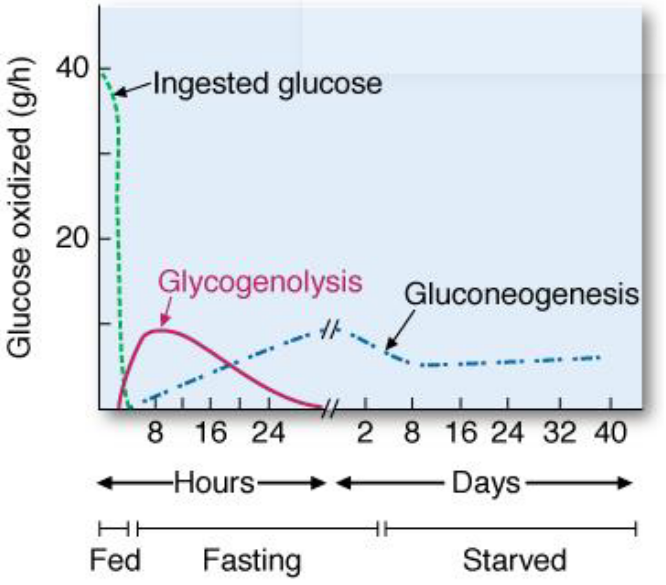
Figure 7.18: Sources of Blood Glucose
During a 12-hour fast, glycogenolysis is the major source of blood glucose. However, at the 16 hour mark, glycogenolysis and gluconeogenesis contribute equally to the maintenance of blood glucose levels.
At the 30 hour mark, liver glycogen stores become substantially depleted - gluconeogenesis is then the primary source of blood glucose.