Paleobiología Esencial: Guía para Estudiantes de Posgrado
2024-12-11
1 Introducción
1.1 Conceptos básicos y contexto histórico
La paleobiología es la rama de la paleontología y de la biología que se ocupa del estudio de los organismos del pasado (entidades paleobiológicas) que conocemos a partir del registro fósil, así como de las relaciones que hubiera habido entre ellos y con su entorno (paleoecología), de la distribución espacial (paleobiogeografía) y de las relaciones filogenéticas que los vinculan (evolución biológica).
El término “fósil” proviene del latín fosillis, que significa “excavado”. Los fósiles son evidencias directas o indirectas de organismos que se encuentran contenidas en la litosfera. Las evidencias directas se refieren a la conservación de un organismo o partes de su cuerpo, lo que nos puede proporcionar una visión detallada de la biología y morfología de los seres vivos del pasado. Por otro lado, las evidencias indirectas se refieren a la preservación de estructuras o huellas de actividad orgánica, que pueden incluir rastros, pistas, o marcas de interacción con el entorno, pero que no involucran la conservación de partes del cuerpo.
Una de las características más destacadas de la paleontología en general, y la paleobiología en particular, es su capacidad única para explorar la dimensión temporal, específicamente el tiempo geológico profundo. El registro fósil proporciona una ventana inigualable hacia el pasado biológico de nuestro planeta, permitiendo a los investigadores rastrear la evolución de la vida a lo largo de millones de años. Esta capacidad de acceder a escalas temporales geológicas profundas es la principal aportación de la paleobiología en el contexto evolutivo. Al preservar evidencia de organismos extintos y sus relaciones con el entorno a lo largo de extensos períodos, el estudio del registro fósil nos permite reconstruir la historia de la vida en la Tierra a una escala de tiempo que ninguna otra disciplina puede igualar.
Es importante destacar que la paleobiología es una disciplina histórica. Las disciplinas históricas se caracterizan por su enfoque en el estudio de eventos y procesos que ocurrieron en el pasado, y lo hacen a través de una variedad de fuentes y metodologías. En el caso de la paleobiología, estas fuentes son los fósiles y sus contextos geológicos. Esta naturaleza histórica está en el centro del debate entre contingencia y determinismo. La visión contingente sostiene que los eventos biológicos y evolutivos dependen de múltiples factores impredecibles, lo que significa que podrían haber sucedido de manera diferente o incluso no haber sucedido. Por otro lado, la perspectiva determinista plantea que estos eventos están regidos por leyes subyacentes o patrones predecibles, lo que implica que el resultado habría sido el mismo, independientemente de las circunstancias específicas.
1.1.1 Los fundadores de la paleobiología
1.1.1.1 Ferenc Nopcsa (1877 - 1933)

Figura 1.1: Baron Nopcsa in Albanian uniform (1915). Imagen disponible bajo licencia Creative Commons CC0.
Ferenc Nopcsa (Fig. 1.1), considerado uno de los fundadores de la Paleobiología, fue un personaje singular en la historia de la ciencia, cuya vida estuvo marcada por una serie de eventos y circunstancias extraordinarias. Nacido en 1877 en Transilvania, en ese momento parte del Imperio Austro-Húngaro, Nopcsa provenía de una distinguida familia de aristócratas húngaros. Su fascinación por la paleontología y la geología se encendió cuando, en 1895, su hermana menor, Ilona, descubrió restos óseos de dinosaurios en la finca familiar en Szentpéterfalva en Săcele, Transilvania. Este descubrimiento impulsó a Nopcsa a inscribirse en la Universidad de Viena para estudiar geología, donde su progreso fue tan notorio que a la temprana edad de 22 años ya comenzó a impartir lecciones. Nopcsa no solo se dedicó al estudio de los reptiles del Mesozoico, sino que también demostró un fuerte interés en la independencia de Albania. A lo largo del tiempo, estableció vínculos con líderes de la resistencia nacionalista albanesa, que luchaba contra el dominio otomano en la región. Nopcsa se convirtió en un apasionado defensor de la causa albanesa, pronunciando discursos inspirados y participando en el contrabando de armas. En 1912, los Estados de los Balcanes unieron sus fuerzas para expulsar a los turcos, un logro que Nopcsa consideró un triunfo significativo para la independencia albanesa. Durante la Primera Guerra Mundial, Nopcsa desempeñó un papel inusual como espía para Austria-Hungría, una faceta de su vida que se suma a la intriga que rodea su historia. Después del conflicto, necesitó encontrar un empleo remunerado y, eventualmente, asumió el cargo de jefe del Instituto Geológico de Hungría, donde pudo seguir contribuyendo al avance de la geología y la paleontología. A pesar de sus logros profesionales, Nopcsa enfrentó dificultades financieras en su vida personal. En un giro trágico, se vio obligado a vender su valiosa colección de fósiles al Museo de Historia Natural de Londres para cubrir sus deudas. Esto marcó el comienzo de una espiral descendente en su vida. En 1933, sumido en una profunda depresión, Nopcsa enfrentó una tragedia aún mayor. Trágicamente, acabó con la vida de su amante, Bayazid Doda, antes de quitarse la vida con un disparo.
Ferenc Nopcsa dejó un legado significativo en la ciencia a través de sus investigaciones pioneras e ideas revolucionarias. A lo largo de su carrera, Nopcsa realizó contribuciones notables que influyeron en diversos campos de estudio. A continuación, destacaremos algunas de sus contribuciones más significativas:
Padre de la paleobiología (paleofisiología): Ferenc Nopcsa es ampliamente reconocido como uno de los fundadores de la paleobiología, aunque en su época él la llamó “paleofisiología”. Su enfoque único implicó explorar lo que se podía inferir sobre la biología de los animales antiguos a partir de los fósiles. Si bien el término “paleobiología” fue acuñado por Othenio Abel en 1911, el trabajo pionero de Nopcsa en este campo estableció las bases para la disciplina.
Origen del vuelo en las aves: Nopcsa desempeñó un papel fundamental en la teorización del origen del vuelo en las aves. Su contribución incluyó la popularización del término “Proavis” (o “Pro-Aves”), aunque William Plane Pycraft lo mencionó por primera vez en 1906. Lo más notable de la aportación de Nopcsa fue la hipótesis de que un ancestro cursorial (corredor) estaba involucrado en la evolución del vuelo en las aves, en contraposición a la idea de un ancestro arbóreo. Su modelo propuso que este “Pro-Avis” habría sido un animal que corría, utilizando las extremidades anteriores emplumadas para acelerar y prolongar sus saltos.
Hipótesis del enanismo insular: Nopcsa también contribuyó significativamente a la comprensión de la evolución en islas a través de su hipótesis del enanismo insular. Sugirió que en ambientes insulares con recursos limitados, los animales tienden a reducir su tamaño a lo largo de las generaciones. Esta teoría, conocida como la regla de la isla, ha sido ampliamente aceptada y ha ayudado a explicar por qué se encuentran especies de menor tamaño en islas comparadas con sus parientes continentales.
Dimorfismo sexual en dinosaurios: Nopcsa fue uno de los primeros científicos en discutir y considerar el dimorfismo sexual en dinosaurios. Su trabajo se centró en la identificación de características anatómicas que podrían indicar diferencias entre machos y hembras dentro de una especie. Aunque su investigación en este campo fue precursora, hoy en día el dimorfismo sexual en dinosaurios es un tema de estudio amplio y continuo.
Exploración geológica de los Balcanes Occidentales: Además de sus contribuciones en paleobiología y paleontología, Nopcsa fue uno de los primeros geólogos en explorar la geología de los Balcanes Occidentales. Su trabajo en esta región ayudó a comprender mejor la estructura geológica y la historia de la Tierra en esta área geográfica específica.
Lectura recomendada
Weishampel, D. B. (2023). Franz Baron Nopcsa: A short life of research in dinosaur evolutionary paleobiology and Albanology. The Anatomical Record, 306(7), 1969-1975.
1.1.1.2 Othenio Abel (1875 - 1946)

Figura 1.2: Othenio Abel (1875–1946) and the cover of Grundzüge der Palaeobiologie der Wirbeltiere. Imagen de Kutschera (2007).
Othenio Abel (Fig. 1.2), un ilustre paleontólogo austrohúngaro nacido en Viena, desempeñó un papel fundamental en el establecimiento de la paleobiología como una disciplina científica. A lo largo de su carrera, Abel dejó una marca perdurable en la ciencia que influyó en el estudio de organismos fósiles y su historia evolutiva. A continuación, exploraremos sus contribuciones más destacadas:
Introducción del Término “Paleobiología”: La contribución más destacada de Othenio Abel a la paleobiología fue la introducción del propio término “paleobiología” en su libro “Grundzüge der Paläobiologie der Wirbeltiere” (Principios de la paleobiología de los vertebrados). En este trabajo, Abel hizo una propuesta audaz al definir la paleobiología como “la rama de las ciencias naturales que pretende dilucidar las adaptaciones de los organismos fósiles y su modo de vida”. Además, agregó que la paleobiología también abordaría “la dilucidación de las relaciones filogenéticas”. Estas ideas revolucionarias estaban en marcado contraste con las corrientes de pensamiento de su época y sentaron las bases de la paleobiología moderna.
El Enfoque Interdisciplinario: Abel no solo acuñó el término “paleobiología”, sino que también abogó por un enfoque interdisciplinario en el estudio de los organismos fósiles. Reconoció que, para comprender plenamente la biología y la evolución de las especies extintas, era esencial combinar métodos y enfoques de diversas disciplinas científicas. Esto incluye la paleontología, la biología evolutiva, la anatomía comparada y la ecología, entre otros campos.
La Contracorriente de su Época: Las ideas audaces de Othenio Abel sobre la paleobiología desafiaron las corrientes de pensamiento prevalecientes en su época. En un momento en que la paleontología se centraba en la descripción y clasificación de fósiles, Abel propuso una visión más amplia que se centraba en la comprensión de la biología y la ecología de los organismos extintos. Esta perspectiva revolucionaria allanó el camino para una comprensión más profunda de la vida del pasado.
Lectura recomendada
Kutschera, U. (2007). Palaeobiology: the origin and evolution of a scientific discipline. Trends in ecology & evolution, 22(4), 172-173.
1.1.2 Relevancia del estudio del contexto histórico
Explorar el contexto histórico de la paleobiología es un ejercicio fundamental para comprender el estado actual del conocimiento en esta disciplina. Este análisis retrospectivo nos recuerda que muchas de las ideas y conceptos que hoy consideramos establecidos no siempre fueron evidentes, invitándonos a cuestionar certezas y a reconocer la naturaleza dinámica de las teorías científicas. A lo largo del tiempo, estas teorías han evolucionado, reflejando no solo los avances técnicos, sino también las influencias culturales y filosóficas que han orientado la investigación científica.
La paleobiología, como cualquier otra rama del conocimiento, ha sido moldeada por el contexto histórico y las creencias predominantes de cada época, lo que demuestra la complejidad inherente a la investigación científica. Como reza el proverbio, “la fe mueve montañas”, y en esta disciplina, las “montañas” de información y conocimiento han sido construidas y transformadas con el tiempo. Comprender este proceso es esencial para apreciar plenamente cómo hemos llegado al conocimiento actual.
Asimismo, es importante destacar que hablar de un progreso absoluto en paleobiología puede resultar problemático. El estudio de la historia de la vida en la Tierra es una obra en constante revisión, y cada nuevo descubrimiento nos obliga a reinterpretar nuestras conclusiones anteriores, reafirmando que la ciencia no avanza de manera lineal, sino a través de un proceso continuo de ajuste y refinamiento.
1.1.3 La fascinación por los fósiles en la prehistoria
El interés por los fósiles se remonta a la prehistoria misma, cuando los primeros seres humanos entraron en contacto con estos restos. A lo largo de milenios, los fósiles no solo estimularon la imaginación de diversas culturas, sino que también encontraron aplicaciones prácticas en múltiples sociedades prehistóricas alrededor del mundo. Existen múltiples ejemplos de cómo los humanos de la prehistoria interactuaron con los fósiles. A continuación, se presentan algunos casos bien documentados.
En Arcy-sur-Cure, una cueva en Borgoña, durante el Paleolítico hace aproximadamente 80,000 años, se ha descubierto la evidencia más antigua de que el ser humano entró en contacto con los fósiles. En esta ubicación se encontró una pequeña colección de gasterópodos y corales fósiles, que se cree fue formada por un grupo de neandertales que ocuparon la cueva. El propósito de estos fósiles en manos de los neandertales sigue siendo una incógnita, pero es probable que fueran utilizados con fines ornamentales, como amuletos, objetos con propiedades mágicas o religiosas. Los fósiles, en este contexto, podrían haber tenido un significado cultural y espiritual para estas primeras culturas humanas.
La conexión entre el ser humano y los fósiles continuó en el Neolítico, hace unos 10,000 años. En Dunstable Down, Inglaterra, se encontró un entierro que destaca por su singularidad. Alrededor del cuerpo de una presunta madre que abraza a su hijo, se colocaron equinoideos irregulares fósiles. Estos fósiles, con su apariencia única, podrían haber tenido algún significado simbólico o ritual en la cultura de la época.
El uso de fósiles no se limitó a Europa. Por ejemplo, en América del Norte, varias culturas prehistóricas utilizaron dientes de tiburón fósiles durante los últimos 10,000 años. Los datos arqueológicos de la región de la Bahía de Chesapeake indican que las culturas nativas recolectaron, modificaron y usaron diferentes variedades de estos dientes. Estos incluían dientes fósiles de tiburones como el gran tiburón blanco (Carcharodon carcharias) y cinco taxones extintos, como el gigantesco megalodón (Otodus megalodon). Los dientes se modificaron de diversas maneras, incluyendo muescas o perforaciones. Aunque muchos probablemente se usaron como puntas de proyectil, cuchillos o herramientas para raspar, algunos presentaban agujeros perforados intencionalmente cerca de las áreas de las raíces, indicando que algunos posiblemente se utilizaron con fines ornamentales, religiosos o como curiosidades.
1.1.4 Fósiles y mitología en la antigüedad
En la Edad Antigua, las mitologías grecolatinas dieron vida a un vasto catálogo de seres mitológicos que combinaban creencias religiosas y observaciones de la naturaleza. Estos seres mitológicos tenían orígenes diversos, desde procesos naturales hasta creaciones divinas, y se convirtieron en parte esencial del imaginario de la antigüedad. Entre los seres mitológicos, encontramos centauros, quimeras, minotauros, hidras, cíclopes, nereidas, sirenas y tritones, que enriquecieron las narrativas de la mitología griega y romana.
La observación de la naturaleza desempeñó un papel fundamental en la creación de estas mitologías. Los antiguos griegos y romanos utilizaban hallazgos fósiles como pruebas tangibles de la existencia de algunos de estos seres mitológicos. Estos vestigios del pasado se integraron en sus mitos y leyendas, y su significado se enriqueció a medida que se tejían relatos más elaborados.
Un ejemplo destacado de esta fusión entre fósiles y mitología se encuentra en la isla de Samos. C. Forsyth Major, alrededor de 1888, descubrió una rica fauna de vertebrados miocénicos en esta isla después de conocer los relatos de Éforo sobre las aneades. Estas aneades eran consideradas como grandes bestias que ataban a las personas en la región y que supuestamente podían destruir la Tierra con el sonido de sus voces. También se basó en los relatos de Plutarco, donde los esqueletos que los griegos encontraron en Samos fueron interpretados como los restos de las amazonas muertas por Dionisio.
La interacción entre los seres humanos y los fósiles no se limitó a la geografía griega. Hace aproximadamente 5,000 años, marineros aqueos que exploraban Sicilia descubrieron fósiles de grandes mamíferos, entre ellos los de proboscidios, que posiblemente eran mastodontes o elefantes enanos. Al observar el tamaño de los huesos, estos marinos pensaron que se trataba de los restos de enormes seres con forma semi-humana a los que llamaron cíclopes, imaginando que tenían un solo ojo. Esta idea surgió debido a la forma del cráneo de los proboscidios, que presentaba una foseta nasal en la región central que fue interpretada como la cavidad ocupada por el único ojo del gigante.
Este asombroso hallazgo y otros similares alimentaron el desarrollo posterior de diversas leyendas sobre la existencia de seres gigantescos. Por ejemplo, en el siglo V, el historiador Tucídides destacó la abundancia de restos de proboscidios en las laderas del Etna, al señalar este lugar como el sitio donde el cíclope Polifemo, hijo del dios Poseidón, devoró a parte de la tripulación del barco de Ulises, de acuerdo a la Odisea de Homero. Las descripciones del tamaño de estos gigantes variaron, desde más de 13 metros, según Plinio, hasta cuatro metros, según Herodoto.
Este vínculo entre los fósiles y la mitología en la Antigüedad ilustra cómo la observación de la naturaleza y la imaginación humana se entrelazaron para crear mitos y leyendas que han perdurado a lo largo de la historia. Estos relatos también han dejado una marca indeleble en la comprensión de la relación entre la ciencia y la cultura en la Antigüedad.
1.1.5 Las primeras ideas griegas sobre los fósiles: cambios en los océanos y la configuración de la Tierra
En la Antigua Grecia, durante los siglos V y VI a.C., algunos pensadores notables como Anaximandro, Jenófanes y Heródoto comenzaron a reflexionar sobre la presencia de fósiles marinos en regiones montañosas. Sus ideas pioneras ofrecieron explicaciones precursoras sobre cómo estos fósiles llegaron a su ubicación actual.
Según sus propuestas, la presencia de fósiles marinos en zonas montañosas sugiere que en el pasado ocurrieron cambios significativos en la configuración o en el nivel de los océanos. En otras palabras, estos pensadores griegos sugirieron que los fósiles de criaturas marinas encontrados en tierras elevadas indicaban que estas áreas alguna vez estuvieron sumergidas bajo el agua. Para ellos, la presencia de restos marinos fosilizados en lugares distantes del mar se convirtió en evidencia de que la Tierra había experimentado transformaciones geológicas a lo largo del tiempo.
Estas primeras ideas, formuladas hace más de dos milenios, anticiparon la comprensión moderna de la geología y la paleontología. A medida que la ciencia avanzó, estas observaciones iniciales se consolidaron en teorías más completas sobre la historia de la Tierra y la vida en ella, marcando el inicio de una investigación científica que sigue evolucionando hasta el día de hoy.
Lectura recomendada
Martín, C. S. (2014). Heródoto (II. 86-88) y el conocimiento anatómico griego. Ágora. Estudos Clássicos em debate, (16), 29-40.
1.1.6 Fósiles en otras culturas: amuletos, talismanes y visiones divinas
El impacto de los fósiles en las creencias y mitologías humanas no se limita a las civilizaciones griegas y romanas. Diversas culturas de todo el mundo también han establecido conexiones fascinantes entre los fósiles y su imaginario colectivo.
En China, la creencia en dragones ha sido una parte integral de su herencia cultural durante siglos. Esta creencia viajó desde Asia hasta Europa, siendo asimilada por griegos y romanos y posteriormente heredada por la cultura occidental. Los chinos han interpretado diversas piezas dentales fósiles como los dientes de dragón. Estos fósiles se convirtieron en objetos de gran valor, utilizados como amuletos, talismanes y, sobre todo, como medicamentos casi milagrosos. La divinidad benefactora que representa el dragón ha llevado a la creencia en los poderes curativos de estos dientes, que han sido utilizados para tratar diversas enfermedades o incluso como afrodisíacos. Actualmente, en muchas partes del mundo, incluyendo China, persisten farmacias chinas que ofrecen estos “dientes de dragón” como remedios tradicionales. Además de los dientes, grandes huesos fosilizados también fueron considerados como los restos de los dragones que no habían encontrado una nube que les permitiera entrar en el cielo. Este vínculo entre los fósiles y la mitología china ilustra cómo la cultura ha influido en la percepción de los fósiles a lo largo de la historia.
Por otro lado, en la antigua Egipto, los sacerdotes utilizaban los amonites, conocidos como cuernos de Amón, con un propósito diferente. Estos fósiles se consideraban elementos que permitían tener premoniciones y visiones divinas durante el sueño. Esta idea se basaba en la similitud en la forma de la concha de estos moluscos con la de los cuernos del carnero, que representaba al dios Amón-Ra. Los sacerdotes egipcios creían que los amonites tenían un vínculo especial con la divinidad y, por lo tanto, les conferían la capacidad de acceder a visiones y conocimiento divino.
Estos ejemplos destacan cómo la fascinación humana por los fósiles trasciende las fronteras culturales y ha dado forma a una diversidad de creencias, rituales y tradiciones en todo el mundo. Los fósiles han sido interpretados de maneras asombrosas y variadas, enriqueciendo la riqueza de las historias humanas y demostrando cómo la ciencia y la cultura se entrelazan de maneras sorprendentes a lo largo de la historia.
1.1.7 La edad del oscurantismo (Edad Media): fósiles como desafío a la Fe y orígenes de mitos
Durante la Edad Media, un período caracterizado por la predominancia de la religión en la vida cotidiana, los fósiles fueron considerados con escepticismo y connotaciones místicas. Para muchos, los fósiles eran simples curiosidades de la naturaleza, mientras que para otros eran trampas puestas por el diablo para poner a prueba la fe de los creyentes. Las creencias de la época, basadas en la teología y la filosofía aristotélica, llevaban a considerar los fósiles como exhalaciones de la tierra a los que les faltó la vis plastica (fuerza vital) o un soplo divino para alcanzar el nivel de seres vivos.
Un ejemplo de estas ideas puede encontrarse en la obra de Plinio el Viejo, quien vivió en el siglo I. Plinio llamó a los dientes fósiles de tiburón glossopetrae, que se traduce como “lenguas petrificadas”, y los interpretó como lenguas petrificadas que caían como lluvia durante los eclipses lunares. También describió los caparazones de erizos de mar como “pequeñas tortugas o huevos de serpiente”. Estos fósiles eran utilizados como amuletos por los druidas para obtener favores de los príncipes. El ámbar, conocido por sus propiedades eléctricas y en griego como electrum, fue descrito por Plinio como “orina de lince petrificada”.
Estas creencias, junto con muchas otras ideas generadas en diferentes culturas y momentos, se transmitieron y evolucionaron a lo largo de la Edad Media, influyendo en la percepción de los fósiles. Por ejemplo, las glossopetrae fueron renombradas como “lenguas de San Pablo”, atribuyéndoles un origen distinto como dientes de serpientes. Se decía que eran lenguas de serpientes que habían mordido al apóstol San Pablo en la isla de Malta, y se les atribuían poderes medicinales y la capacidad de absorber venenos. En contraste, fueron uno de los primeros fósiles a partir de los cuales se postula el origen orgánico de estos restos. Los amonites se pensaba que eran serpientes petrificadas. Para aumentar su valor y hacerlos más atractivos para los compradores, los falsificadores solían esculpir cabezas de serpiente en estos fósiles para hacerlos parecer aún más auténticos. Estas “serpientes petrificadas” eran considerados objetos especiales y a menudo se utilizaban como amuletos o elementos decorativos. Un ejemplo curioso relacionado con los amonites se encuentra en algunas culturas prehispánicas de México, donde estos fósiles fueron interpretados como “rayos de sol petrificados”. Las ostras del género Gryphae, con su apariencia peculiar, fueron consideradas por algunas personas de la Edad Media como “pezuñas perdidas por el diablo”. Durante este periodo, la gente creía en la existencia del unicornio, que era plasmado en historias escritas, pinturas y esculturas. Este ser mitológico se originó muy posiblemente a partir de una interpretación errónea de varios animales, como el rinoceronte o el órix. El cuerno del unicornio se creía que tenía propiedades medicinales y afrodisíacas. Estas creencias se relacionaron con las cualidades atribuidas al cuerno de rinoceronte en algunas culturas de Asia, Medio Oriente y África. Esta percepción del cuerno del unicornio como un elemento valioso impulsó una búsqueda frenética de estas piezas en el mercado, lo que a menudo llevó a la comercialización de otros objetos con forma similar. En lugar de auténticos cuernos de unicornio, en los mercados de la época, era posible encontrar defensas de proboscidios (como colmillos de elefantes) y los incisivos de narvales actuales y fósiles. Estos últimos, con su característica forma espiralada, eran particularmente valorados y a menudo se representaban como cuernos de unicornio en el arte del medioevo y del Renacimiento. Otro ejemplo es la interpretación errónea de fósiles como restos de gigantes, lo que llevó a la creación de mitos como la leyenda de Teotobocus, un rey germánico derrotado por el ejército romano. Sus supuestos restos óseos eran en realidad huesos de mastodontes. Además, varios héroes cristianos se enfrentaron a seres malignos y gigantes, como dragones, en leyendas vinculadas a la religión. Esos seres mitológicos se creían apoyados por la aparición de esqueletos fosilizados de diferentes animales, como osos del Pleistoceno.
1.1.8 Fósiles y leyendas en las antiguas culturas de América
En América prehispánica, varias culturas desarrollaron mitos y leyendas basados en restos fosilizados de plantas y animales. Al igual que en el Viejo Mundo, el hallazgo de grandes huesos o troncos fosilizados generó la creencia en gigantes en algunas culturas indígenas. El pueblo Navajo creía que los troncos petrificados encontrados en Arizona eran los huesos del gigante Yetso. Según su mito, sus antepasados habían tenido que matar a este gigante para poder habitar esas tierras. Otros antiguos habitantes de la región, como los Anasazi, interpretaron esos mismos troncos como enormes flechas o armas rotas utilizadas durante batallas entre gigantes. Fray Juan de Torquemada describe en su obra “Monarquía indiana” las creencias de los toltecas sobre la existencia de grandes hombres llamados Quinametin, que poblaron la región antes de la llegada de los hombres normales. La leyenda tolteca afirmaba que estos gigantes llegaron por mar, construyeron grandes edificaciones y que fueron castigados con un fuego divino debido a sus pecados. Los toltecas presentaban huesos fosilizados de grandes mamíferos terciarios como prueba de la existencia de estos gigantes. Fray Juan de Torquemada, con el propósito de respaldar la idea de la existencia de gigantes en América, relató entrevistas con europeos que afirmaron haber visto enormes huesos, cráneos, mandíbulas y piezas dentales de estos gigantes. La crónica de Bernal Díaz del Castillo, titulada “Historia verdadera de la conquista de la Nueva España”, amplía la leyenda tolteca al relatar que los tlaxcaltecas presentaron a Hernán Cortés un inmenso hueso como prueba de la existencia de gigantes contra los cuales sus antepasados habían luchado para conquistar la región. Según la leyenda tlaxcalteca, para derrotar a estos gigantes, sus ancestros habían aprovechado la glotonería y la afición por las bebidas alcohólicas de los gigantes. Organizaron un festín y esperaron a que la embriaguez y la comida hicieran dormir a los gigantes, momento en el que los mataron. El hueso presentado a los españoles, según el relato de Díaz del Castillo, persuadió a Cortés acerca de la existencia de gigantes en América, lo que lo llevó a decidir enviarlo a España como evidencia. La cultura Totonaca, en la región que actualmente abarca Guatemala y México, estaba familiarizada con los fósiles de amonitas, a los que llamaban “solecitos”. Estos fósiles, comunes en las rocas de la zona, fueron relacionados con el Sol. Los Totonacas creían que los “solecitos” eran reflejos o rayos de luz solar petrificados, dejados por sus dioses como una señal de su existencia. Una de las leyendas más conocidas de las culturas prehispánicas en la región de Papantla (hoy en México) está relacionada con el ritual de los Voladores de Papantla. En este ritual, los “voladores” realizan un vuelo imaginario que representa al Sol y a la lluvia al caer sobre la tierra, al tiempo que veneran a las deidades correspondientes. Los fósiles de amonitas eran familiares para la cultura totonaca y se relacionaban con el Sol, ya que los consideraban reflejos o rayos de luz solar petrificados, dejados por sus dioses como una señal de su existencia. En este ritual, utilizan un penacho construido con plumas dispuestas helicoidalmente, que reproduce los destellos iridiscentes del Sol y la forma de las amonitas.
Lectura recomendada
Tovar, F. S., & Barroso, S. A. Q. (2014). Mitos y leyendas sobre los fósiles. Ciencia, 8-16.
1.1.9 Pensamiento renacentista y las interpretaciones de los fósiles
En el período del Renacimiento, se retomaron las ideas de pensadores griegos de los siglos V y VI antes de Cristo, como Anaximandro, Jenófanes o Heródoto, que sostenían que el hallazgo de fósiles marinos en zonas montañosas evidenciaba cambios en la configuración o el nivel de los océanos en el pasado. A pesar de estas ideas, las interpretaciones sobre los fósiles seguían estando influenciadas por creencias religiosas, particularmente la noción del diluvio universal como explicación. La idea de que el supuesto diluvio bíblico podía explicar la presencia de restos de organismos marinos en zonas continentales fue formulada por primera vez en el siglo XIII por un monje italiano llamado Ristoro de Arezzo. Esta idea fue posteriormente desarrollada por otros pensadores en el Renacimiento. Un evento importante en este contexto ocurrió en 1725, cuando el médico naturalista ruso Jacob Scheuchzer presentó al mundo su hallazgo conocido como el Homo diluvii testis, al que describió como los restos de un hombre que fue testigo del diluvio (Fig. 1.3). Sin embargo, en el siglo XIX, el anatomista francés George Cuvier analizó el esqueleto y demostró que en realidad se trataba del fósil de una salamandra de gran tamaño.

Figura 1.3: Especimen de Andrias scheuchzeri que inspiró las ideas de Jacob Scheuchzer sobre el Homo diluvii testis. Imagen disponible bajo licencia Creative Commons CC0.
En el siglo XVI, se comenzó a observar un estudio más sistemático de los fósiles, sin necesariamente atender a su parecido con entidades orgánicas. Fue en esta época que se realizaron las primeras representaciones de fósiles en libros impresos gracias a la recién inventada imprenta y técnicas de reproducción seriada lo que permitió una difusión más amplia del conocimiento y contribuyó significativamente al estudio de los fósiles y la paleontología.
En 1557, Christoph Encellius, un sacerdote alemán y geólogo de renombre, ilustró un fósil de bivalvo con forma de vieira en su libro “De Re Metallaci” (Sobre los metales) (Fig. 1.4A). Encellius agrupó los fósiles con los minerales, considerándolos eventos naturales que coincidentemente se parecían a las formas de los seres vivos.
Además, otros científicos de la época, como Georgius Agricola, médico y farmacéutico en Sajonia, contribuyeron al estudio de los fósiles. Agricola escribió “De Natura Fossilium” en 1546, donde propuso una clasificación de los fósiles basada en propiedades físicas como la dureza, el lustre y la facilidad para pulirlos. También creía que los fósiles se formaban por la acción de un fluido cementante que circulaba dentro de la Tierra.
Otro científico destacado de esa época fue Conrad Geissner, quien en 1565 publicó “Rerum Fossilium Lapidum et Gemmarum” la primera monografía ilustrada sobre fósiles. Geissner clasificó los fósiles en 15 clases, basándose en su forma o el material. Algunas de estas clases incluían objetos como plantas, partes de animales, objetos marinos y formas geométricas. Sus descripciones no solo se limitaban a las características físicas, sino que también incluían opiniones de autores previos, el significado y origen de los nombres y una relación de las propiedades medicinales de los fósiles. Geissner comparó el dibujo de Encellius con otro de Guillame Rondelet que representaba un molusco vivo (Fig. 1.4B). Geissner se dio cuenta de que Encellius había dibujado un caparazón fosilizado de lo que alguna vez pudo ser un organismo vivo. El trabajo de Geissner marcó un paso importante hacia el pensamiento moderno y la comprensión de que muchas formas petrificadas eran, de hecho, restos de organismos que alguna vez estuvieron vivos. En su libro de 1565 “Sobre objetos fósiles,” Geissner comparó las formas muy similares de dos equinodermos, un erizo de mar moderno y un erizo irregular petrificado, concluyendo que este último debía haber sido una forma viva en el pasado y, por lo tanto, un resto fósil de equinodermo.
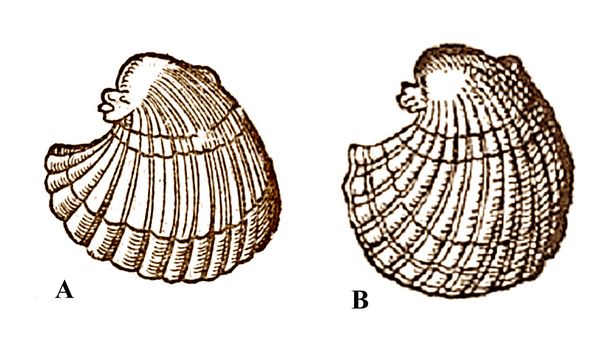
Figura 1.4: Comparación entre el bivalvo fósil ilustrado por Encellius en 1557 (A) y el bivalvo actual ilustrado por Rondelet en 1558 (B), que llevó a Geissner a pensar sobre el posible origen orgánico de los fósiles. © athenaclub.
Hacia finales del siglo XVI y principios del XVII, Italia se convirtió en el centro de interés en torno a los fósiles, destacando cuatro grandes colecciones en Bolonia, Verona, Nápoles y Roma (Vaticano). Cada una de estas colecciones estaba respaldada por naturalistas como Ulisse Aldrovandi, Francesco Calceolari, Ferrante Imperato y Michele Mercati, respectivamente.
1.1.10 El cambio de paradigma en la paleontología del siglo XVII

Figura 1.5: Retrato de Niels Stensen. Imagen disponible bajo licencia Creative Commons CC0.
En el siglo XVII, se produjo un avance significativo en la interpretación de los fósiles. Niels Stensen (1638-1686) (Fig. 1.5), un naturalista danés, desempeñó un papel fundamental en este sentido realizando, en 1667, la primera interpretación documentada sobre el origen orgánico de los fósiles a partir de un grabado del catálogo inédito de Michele Mercati. El método científico, desarrollado en este siglo por figuras como Galileo Galilei, fue aplicado por Stensen a la paleontología.
Para respaldar su interpretación, Stensen llevó a cabo la disección de la cabeza y los dientes de un gran tiburón blanco procedente de Livorno, Italia (Fig. 1.6). Durante su examen, notó la sorprendente similitud entre los dientes de tiburón actuales y las glossopetrae de Malta. Además, observó la presencia de signos de descomposición en estos últimos. Basándose en estas observaciones, Stensen propuso que los fósiles tenían un origen orgánico. En su obra Canis carchariae dissectum caput (cabeza disecada de Canis carchariae) donde publicó estos resultados, afirmó:
“Para concluir esta digresión, usaré algunas de las deducciones que he realizado sobre las grandes piedras en forma de lengua. Que se asemejan a los dientes de tiburón es claramente evidente por su forma, ya que la superficie, el borde y la base son muy similares a las partes correspondientes de los dientes. Si creemos en los informes, nuevas islas continúan surgiendo en medio del mar, y quién sabe cuál fue la primera cuna de Malta. Quizás, en tiempos anteriores, cuando estaba cubierta por el mar, proporcionaba refugio a los tiburones, cuyos dientes quedaron enterrados en el fondo marino. Sin embargo, tras un cambio, debido a la actividad subterránea, los dientes se encontraron en medio de la isla.” En 1668, publicó su obra maestra titulada De solido intra solidum naturaliter contento dissertationis prodromus (Discurso preliminar de una disertación sobre los cuerpos sólidos contenidos de manera natural en un sólido). En esta obra, presentó su teoría sobre el origen orgánico de los fósiles y desarrolló la Ley de superposición de estratos, que se convirtió en un principio fundamental en la geología.
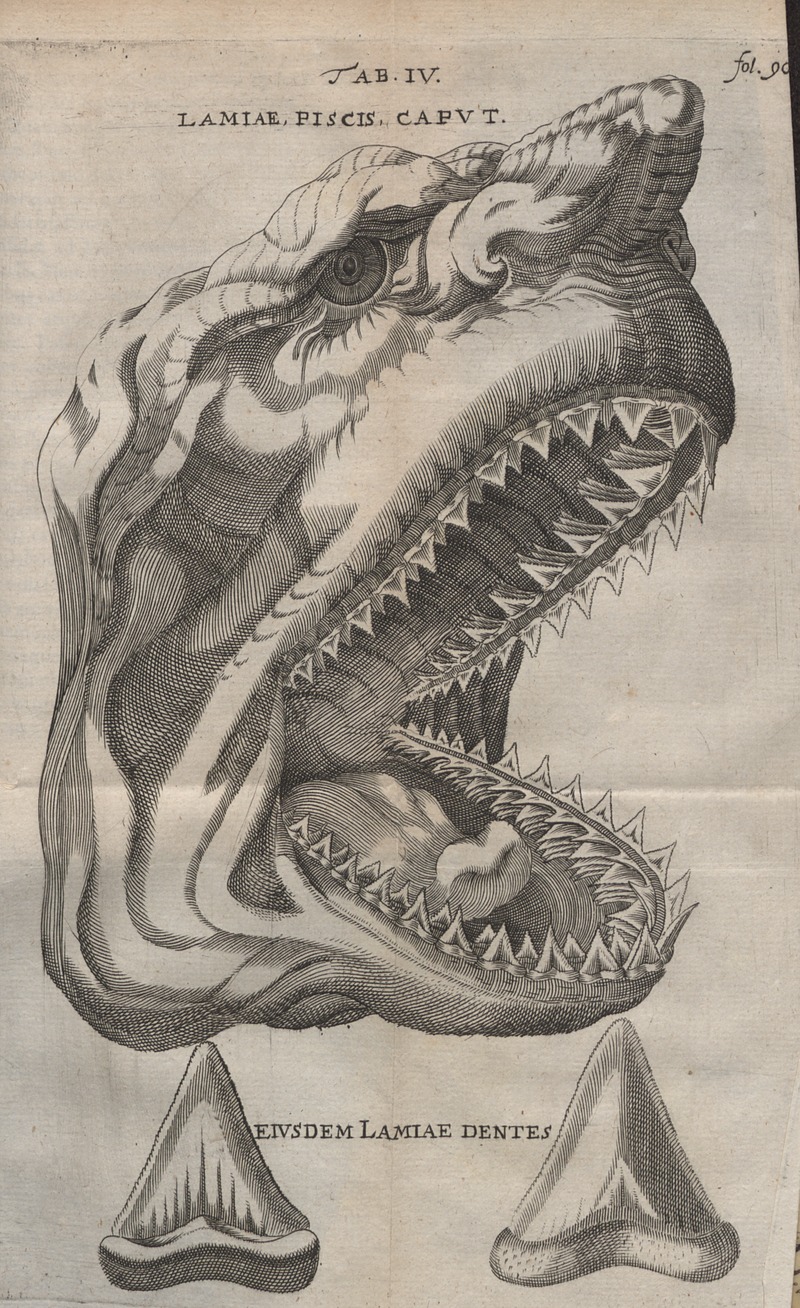
Figura 1.6: Ilustración del artículo de Stensen de 1667 comparando los dientes de una cabeza de tiburón blanco con un diente fósil de Megalodon. Imagen disponible bajo licencia Creative Commons CC0.
La Ley de superposición de estratos establecía que, si se aceptaba el origen orgánico de los fósiles, los materiales rocosos que los contenían debieron haber presentado inicialmente un estado no consolidado. Estos materiales pudieron enterrar los restos de los organismos y sus actividades. Con el tiempo, estos materiales se endurecieron y transformaron en rocas sedimentarias, que se apilaron en capas o estratos. Los estratos más antiguos se encuentran en la parte inferior, mientras que los más modernos están en la parte superior. Stensen también propuso la idea de “agua petrificante” para explicar el aspecto petrificado de los fósiles, respaldado en la evidencia de mineralizaciones rápidas en minas. Esta idea se basó en la creencia en la teoría corpuscular de la materia, que permitía que ciertas sustancias penetren en los materiales y modifiquen su estructura interna. Los alquimistas de la época intentaron transmutar cualquier metal en oro, basándose en esta idea.
Lectura recomendada
Sequeiros, L. (2002). Raíces de la Geología. Nicolás Steno, los estratos y el diluvio universal, Las. Enseñanza de las Ciencias de la Tierra, 10(3), 217-242.
1.1.11 El siglo XVII en Inglaterra
Durante el siglo XVII en Inglaterra, hubo un progreso significativo en el entendimiento de los fósiles y su origen. Este período fue testigo de debates apasionados sobre la naturaleza de los fósiles y su relación con la historia de la Tierra. La obra de Stensen fue traducida al inglés por Henry Oldenburg (Secretario de la Royal Society).

Figura 1.7: Retrato de Robert Hooke. Imagen disponible bajo licencia Creative Commons CC0.
Uno de los puntos de referencia clave de esta época fue Robert Hooke (1635-1703) (presidente de la Royal Society) (Fig. 1.7), cuyo legado dejó una huella indeleble en la paleobiología. En 1665, Hooke publicó su obra maestra, “Micrographia”, que revolucionó el mundo de la ciencia. En este libro, Hooke presentó al mundo una visión fascinante del mundo microscópico. Sus ilustraciones detalladas y precisas, que incluyeron células, término que él mismo acuñó, permitieron a los científicos observar plantas, animales y fósiles a través del microscopio por primera vez.
Hooke no se limitó a la investigación microscópica, sino que también incursionó en el estudio de los fósiles. Fue el primero en examinar estos vestigios antiguos bajo el microscopio. Lo que descubrió fue asombroso. Al comparar la madera petrificada con la madera en descomposición, Hooke llegó a la conclusión de que la petrificación ocurría cuando el agua depositaba minerales en la madera, convirtiéndola en piedra (Fig. 1.8). Este mismo proceso, según sus observaciones, se aplicaba a las conchas fósiles. Para él, las similitudes entre las conchas vivas y las conchas fósiles no eran fruto del azar, sino de la acción de algún tipo de barro, arcilla o agua petrificante.
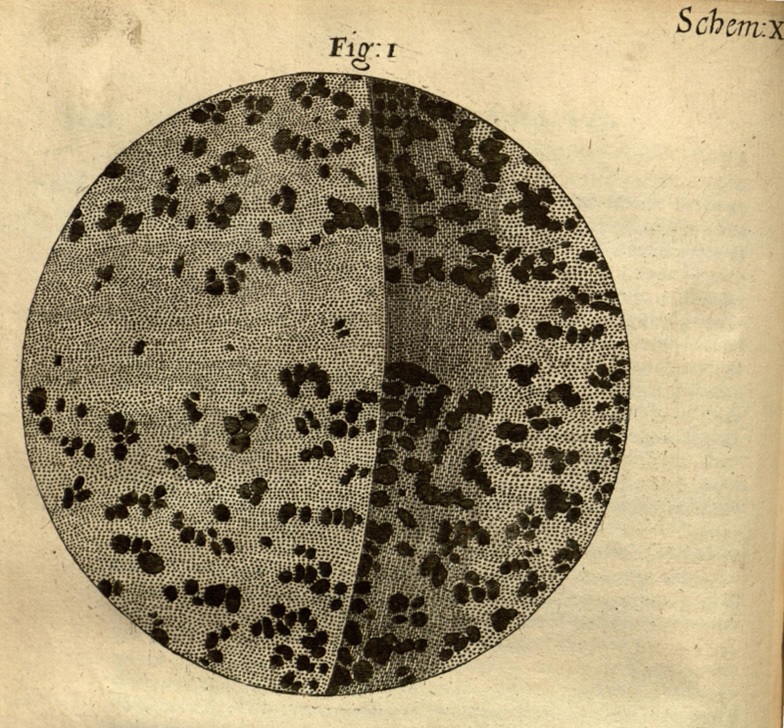
Figura 1.8: Ilustración de madera petrificada (xilópalo) observada bajo el microscopio por Robert Hooke. © biodiversitylibrary.
El debate sobre los fósiles y su origen orgánico no se limitó a Hooke. Otros científicos británicos, como Martin Lister, John Ray y John Woodward, también contribuyeron con sus puntos de vista.
Martin Lister (c. 1638-1712), médico londinense, escribió un libro sobre moluscos vivientes en el que observó que la mayoría de los fósiles británicos no guardaban gran parecido con los animales actuales. Al no aceptar la idea de la extinción de especies, Lister rechazaba el origen orgánico de los fósiles. Dos observaciones reforzaban sus sospechas: muchos de los fósiles eran solo impresiones sin indicios claros de las conchas originales, y ciertos tipos de roca parecían formar estructuras que imitaban conchas específicas.
Otro gran naturalista, John Ray (1627-1705), se mostraba más indeciso en sus conclusiones. Consideraba ambos puntos de vista: por un lado, le parecía increíble que las similitudes entre conchas vivientes y fósiles, incluso a nivel microscópico, pudieran ser producto del azar. Sin embargo, la extinción de organismos como los ammonites sugería una imperfección en la Creación divina, lo cual también le resultaba inconcebible. Ray propuso que quizás las especies fósiles no estuvieran realmente extintas, sino que habitaran en océanos aún inexplorados, aunque esta idea no lo convencía del todo. Además, Ray enfrentaba el problema de la ubicación de los fósiles. Aunque se aceptaba comúnmente que el Diluvio bíblico era el único evento que había alterado la superficie de la Tierra desde su creación, Ray sabía que los fósiles no estaban dispersos en la superficie terrestre, sino incrustados en capas de roca. Esto dificultaba explicar racionalmente cómo una inundación podría haber sido lo suficientemente poderosa como para cubrir incluso los Alpes
John Woodward (1665-1728), uno de los grandes coleccionistas de su época, tenía opiniones bien definidas sobre el tema. En su obra “Essay towards a Natural History of the Earth” (1695), abordó muchas de las incógnitas existentes en torno a los fósiles. Sostenía que las conchas y huesos fósiles eran restos de animales antidiluvianos que, junto con los materiales de la superficie terrestre, habían sido agitados por el Diluvio y quedaron organizados en capas según sus densidades. Además, aceptó la idea de Ray de que esas formas extrañas podrían encontrarse vivas en algún momento futuro.
En conclusión, el siglo XVII en Inglaterra marcó una época de cuestionamiento y exploración en la paleobiología y la paleontología. Desde las revolucionarias observaciones microscópicas de Hooke hasta los debates sobre la extinción y el Diluvio, los científicos de la época sentaron las bases para el entendimiento moderno de los fósiles y su relación con la historia de la Tierra.
1.1.12 El siglo XVIII: cambios en la perspectiva de los fósiles y la influencia de la teología natural
Durante el siglo XVIII el debate que había dominado gran parte del siglo anterior comenzó a perder fuerza en Inglaterra, y surgieron nuevas perspectivas. En el contexto del protestantismo inglés, la teología natural se convirtió en un enfoque ideológico importante. Este enfoque buscaba encontrar evidencia de la existencia de Dios sin recurrir a revelaciones sobrenaturales. La idea de especies extintas planteaba un desafío teológico, ya que implicaba una imperfección en la creación divina, un concepto que generaba debate y reflexión entre los naturalistas. A pesar de los avances en la comprensión de los fósiles, muchos naturalistas todavía sostenían la idea de que la distribución de los fósiles se debía a la acción de un diluvio único. Esta perspectiva se mantuvo arraigada en parte debido a razones teológicas y a la búsqueda de la armonía entre la ciencia y la religión. A mediados del siglo XVIII, hubo un consenso en crecimiento sobre el origen orgánico de los fósiles. Esto representó un cambio significativo en la perspectiva sobre estos restos.
En 1735, Carl Linnaeus, un científico, naturalista, botánico y zoólogo sueco, publicó su obra “Systema Natura”. Linnaeus es considerado el creador de la clasificación de los seres vivos, también conocida como taxonomía. Introdujo un sistema de nomenclatura binomial en el que cada especie se identificaba mediante dos términos (epítetos): el primero, con la letra inicial en mayúscula, representaba el género, y el segundo, en minúscula, el nombre específico de la especie. Este sistema de nomenclatura se convirtió en un estándar en la biología y sigue siendo fundamental en la actualidad.
1.1.13 Avances a finales del siglo XVIII: teoría del origen de la Tierra y del origen orgánico de los fósiles
A medida que el siglo XVIII llegaba a su conclusión, se desarrollaron ideas revolucionarias sobre el origen de la Tierra y la naturaleza de los fósiles, basadas en las contribuciones de pensadores como René Descartes, Niels Stensen y Robert Hooke. Una de las ideas destacadas a finales del siglo XVIII postulaba que la Tierra se originó a partir de un estado incandescente o fundido. Esta perspectiva formulaba que la Tierra se formó a partir de materiales en fusión y experimentó un proceso de enfriamiento y solidificación.
Se propuso una clasificación de los materiales terrestres en tres categorías distintas:
Materiales sin fósiles previos al Diluvio (primario): Estos materiales se consideraban anteriores a cualquier evento catastrófico, como un diluvio, y, por lo tanto, no contenían fósiles.
Materiales del diluvio serían fosilíferos (secundario): Se sostenía que los materiales depositados durante un diluvio único contenían fósiles.
Materiales aluviales no consolidados (terciarios): Esta categoría se refería a materiales más recientes depositados por acción de ríos y corrientes, que aún no se habían endurecido en rocas consolidadas.
La idea de que la Tierra debía tener una edad mucho mayor comenzó a ganar aceptación. El Conde de Buffon, Georges-Louis Leclerc (1707-1788) (Fig. 1.9), propuso que la Tierra podría tener decenas o incluso cientos de miles de años de antigüedad. Esta perspectiva marcó un alejamiento significativo de la concepción tradicional de una Tierra joven y tuvo un impacto profundo en la forma en que se entendía la historia de nuestro planeta. Como anécdota, James Ussher, un erudito irlandés del siglo XVII y arzobispo de Armagh, publicó su obra “Annales Veteris Testamenti” (Anales del Antiguo Testamento) en 1650 donde argumentaba que la creación ocurrió el 23 de octubre del año 4004 a.C. a las 9:00 a.m. Esta cronología se basó en la genealogía y los eventos relatados en el Antiguo Testamento de la Biblia.

Figura 1.9: Retrato del Conde de Buffon. Imagen disponible bajo licencia Creative Commons CC0.
1.1.14 Siglo XVIII y principios del siglo XIX: debates clave en la paleontología y geología
El período que abarca el siglo XVIII y los primeros años del siglo XIX fue testigo de debates fundamentales en las disciplinas de paleontología y geología. Estos debates se centraron en varias posiciones opuestas que influyeron en la comprensión de la historia de la Tierra y de la vida en ella. A continuación, exploraremos los principales debates y divisiones conceptuales que caracterizaron esta época:
Catastrofismo vs. Uniformismo
Catastrofismo: Esta perspectiva sostenía que los cambios en la Tierra, tanto geológicos como biológicos, se debían a eventos catastróficos repentinos, como diluvios o erupciones volcánicas masivas. Los catastrofistas creían en la idea de que la Tierra experimentaba eventos abruptos que tenían un impacto significativo en la vida y en la geología.
Uniformismo: Por otro lado, los uniformistas argumentaban que los cambios en la Tierra y en la vida se producían a lo largo de períodos extremadamente largos y a través de procesos graduales y uniformes. Esta perspectiva sugería que los fenómenos geológicos y biológicos se explicaban mediante causas naturales y procesos lentos y constantes.
Fijismo vs. Transformismo
Fijismo: Los fijistas sostenían la creencia en la inmutabilidad de las especies. Argumentaban que las especies eran invariables y no cambiaban con el tiempo.
Transformismo: Los transformistas defendían la idea de que las especies podían cambiar y evolucionar con el tiempo. Esta perspectiva marcó un paso crucial hacia la teoría de la evolución.
Direccional vs. Estacionario
Direccional: Los defensores de la dirección sostenían que la historia de la Tierra y de la vida se caracterizaba por un progreso continuo y un desarrollo lineal hacia estados más avanzados.
Estacionario: Por otro lado, los partidarios de la estacionariedad argumentaban que la historia de la Tierra y de la vida era cíclica y que no mostraba un progreso lineal sostenido.
Ambientalista vs. Internalista
Ambientalista: Los ambientalistas enfatizaban la influencia del entorno y del clima en la evolución y en los eventos geológicos. Argumentaban que los cambios ambientales impulsaban la evolución y la extinción de especies.
Internalista: Los internalistas, por otro lado, consideraban factores internos, como la lucha por la supervivencia y la competencia entre especies, como motores principales de la evolución y los cambios en la biodiversidad.
Gradual vs. Saltacionista
Gradual: Los gradualistas sostenían que los cambios en la Tierra y en la vida se producían de manera gradual a lo largo del tiempo. Creían en procesos evolutivos lentos y constantes.
Saltacionista: Los saltacionistas argumentaban que los cambios ocurrían de manera brusca y a través de saltos evolutivos, lo que implicaba cambios significativos en períodos de tiempo relativamente cortos.
Durante este período, se establecieron divisiones mayores en la historia de la Tierra y de la vida. Se identificaron las Eras Paleozoica, Mesozoica y Cenozoica, junto con sus subdivisiones principales. Estas divisiones proporcionaron un marco temporal que permitía a los científicos comprender mejor la historia geológica y biológica de la Tierra. Un hito importante de esta época fue el reconocimiento de los fósiles como marcadores temporales clave. Los fósiles se convirtieron en herramientas fundamentales para determinar la edad de las rocas y para establecer la secuencia de eventos en la historia de la Tierra. Este reconocimiento sentó las bases para futuros avances en la paleontología y la geología.
En resumen, el siglo XVIII y principios del siglo XIX fueron testigos de debates cruciales que dieron forma a la comprensión de la Tierra y de la vida en ella. Estos debates plantearon cuestiones fundamentales sobre el cambio, la evolución y la naturaleza misma de nuestro planeta y sus habitantes, y proporcionaron la base para desarrollos posteriores en estas disciplinas científicas.
1.1.15 Siglo XVIII y principios del siglo XIX en París
1.1.15.1 La extinción: los aportes de Cuvier y Brongniart al conocimiento paleontológico

Figura 1.10: Retrato de Georges Cuvier. Imagen disponible bajo licencia Creative Commons CC0.
Este período en París fue testigo de la destacada figura de Georges Cuvier (1769-1832) (Fig. 1.10), quien realizó contribuciones significativas a la comprensión de la extinción y la paleontología. Cuvier, que se desempeñó como profesor de Anatomía en el Musée National d’Historie Naturelle, realizó investigaciones pioneras que marcaron un punto de inflexión en la concepción de la historia de la vida en la Tierra.
Una de las contribuciones más notables de Cuvier fue su reconocimiento de la extinción de especies. Su estudio se centró en la comparación de los esqueletos de elefantes africanos y asiáticos, revelando diferencias que justificaban la clasificación de estas especies en categorías separadas. Al examinar estos restos, Cuvier también observó que tanto los elefantes actuales como los mamuts y mastodontes extintos pertenecían al mismo grupo de los elefantes. Estas observaciones proporcionaron evidencia concluyente de la existencia previa de especies bien conocidas que ya no estaban presentes en la Tierra. Cuvier demostró que los elefantes actuales no eran simplemente una variante de los mamuts, sino especies distintas.
Cuvier planteó la idea de que la extinción de las especies estaba relacionada con causas catastróficas. En su visión, la Tierra había experimentado una “revolución del globo” repentina y generalizada, que estaba vinculada a eventos catastróficos. Este concepto fue influido por el contexto de la Revolución Francesa, y Cuvier lo aplicó a la naturaleza para explicar la desaparición de especies. Una de las causas catastróficas que propuso fue el aumento del nivel del mar, que consideraba suficientemente rápido como para ahogar a las especies antes de que pudieran desplazarse y sobrevivir. Esta hipótesis planteó la posibilidad de que cambios ambientales drásticos pudieran haber desempeñado un papel en la extinción de especies.
Cuvier no se limitó a sus investigaciones sobre elefantes y mamuts. Durante su carrera, describió los restos de una diversidad de vertebrados extintos. Su trabajo incluyó la identificación y descripción de especies como el perezoso gigante de América del Sur, el mastodonte de América del Norte, un hipopótamo, rinocerontes y muchos otros. Cuvier contribuyó significativamente a la paleontología al ampliar la lista de especies fósiles conocidas y proporcionar una comprensión más profunda de la extinción.
Junto con Alexandre Brongniart, Georges Cuvier constató que las formaciones geológicas superpuestas diferían en su contenido fósil, lo que indicaba cambios en la fauna y la flora a lo largo del tiempo. Esta observación lo llevó a proponer la idea de que las especies extintas eran reemplazadas por otras que, aunque no idénticas, eran funcionalmente similares en un proceso de extinción y originación. Cuvier reconoció que para explicar la aparición de nuevas especies era necesario comprender qué mecanismos impulsaban la formación de estas. Las ideas de Cuvier sientan las bases para la noción de creaciones sucesivas, que sostiene que a medida que las especies se extinguían, eran reemplazadas por otras que se adaptaban a las nuevas condiciones del entorno. Este concepto se convirtió en un fundamento para comprender cómo la biodiversidad y la vida en la Tierra evolucionaban con el tiempo.
Adolphe Brongniart, hijo de Alexandre Brongniart, desarrolló y amplió las ideas de Cuvier. En su obra “Histoire des Végétaux Fossiles” (Historia de las Plantas Fósiles) publicada en 1828, Brongniart distinguió cuatro floras sucesivas en el registro fósil. Estas floras representaban diferentes etapas en la evolución de las plantas, desde las criptógamas del Carbonífero hasta las gimnospermas del Mesozoico y finalmente las angiospermas del Terciario, que incluyen la diversidad de plantas actuales. Brongniart relacionó esta progresión con la disminución gradual de los niveles de dióxido de carbono atmosférico y consideró los efectos secundarios de los cambios climáticos y eustáticos en la Tierra.
1.1.15.2 La herencia de los caracteres adquiridos: Lamarck y su impacto en la biología
La idea del cambio biológico es un concepto que ha estado presente en la historia de la humanidad desde tiempos remotos y ha encontrado eco en diversas culturas. Incluso en la antigua Grecia, filósofos como Anaximandro ya especulaban sobre los orígenes de la vida y planteaban la hipótesis de que los animales acuáticos podrían haber dado origen a los terrestres. Esta idea temprana de la transición de formas de vida influyó en futuras concepciones evolutivas. En el Lejano Oriente, antiguos pensadores chinos, como Zhuangzi, sostenían que las formas de vida poseían una habilidad innata para adaptarse a su entorno. Esta noción subrayaba la idea de que los seres vivos eran capaces de cambiar y evolucionar en respuesta a desafíos ambientales. En contraste, los filósofos taoístas en China negaban la inmutabilidad de las especies, abriendo la puerta a la idea de que los organismos podían cambiar con el tiempo. La sociedad musulmana también contribuyó a la evolución del pensamiento evolutivo. En el siglo IX, el erudito Al-Jahiz presentó argumentos sobre la variabilidad de las especies y la selección natural, anticipando algunas ideas clave de la teoría de la evolución. Siglos después, en el siglo XIII, Nasir al-Din al-Tusi exploró conceptos evolutivos en su trabajo. Estos antecedentes históricos muestran que la noción de la evolución biológica ha estado presente en diferentes momentos y lugares, sentando las bases para la teoría moderna que comenzaría a cristalizar en los siglos XVIII y XIX.
Jean Baptiste Lamarck (1744-1829) (Fig. 1.11) fue un precursor en el campo de la biología evolutiva, cuyas ideas representan un hito importante en la comprensión de cómo cambian las formas de vida a lo largo del tiempo. A pesar de que su enfoque sobre la evolución se desarrolló en un contexto histórico en el que la generación espontánea era ampliamente aceptada, Lamarck presentó conceptos audaces y revolucionarios que allanaron el camino para la comprensión moderna del cambio biológico.
Figura 1.11: Retrato de Jean Baptiste Lamarck. Imagen disponible bajo licencia Creative Commons CC0.
En la perspectiva de Lamarck, los animales avanzaban a lo largo de una “escalera de la vida”, donde cada peldaño representaba un nivel creciente de complejidad y organización. Creía que los organismos modificaban su forma en respuesta a las presiones ambientales y a la necesidad de adaptarse. Esta noción fue una de las primeras hipótesis evolutivas que planteaba que los cambios en el entorno podían dar lugar a modificaciones en las características de los organismos. Un componente fundamental de la concepción de Lamarck era la idea de que los organismos adquirían rasgos durante su vida y luego transmitían dichos rasgos a su descendencia (herencia de los caracteres adquiridos). Por ejemplo, si un organismo desarrollaba músculos más fuertes debido a su actividad, Lamarck sostenía que sus descendientes heredarían estos músculos fortalecidos.
A pesar de que la propuesta de Lamarck fue innovadora en su enfoque sobre la evolución, fue desplazada posteriormente por el concepto de la selección natural de Charles Darwin. No obstante, Lamarck realizó aportes significativos a la biología en su conjunto, incluyendo la distinción clara entre el mundo orgánico y el inorgánico, la concepción de la organización de los seres vivos y una clasificación revolucionaria de los animales según su complejidad. Además, formuló la primera explicación del cambio biológico, sentando las bases para futuros avances en el campo de la biología evolutiva.
1.1.15.3 Del fijismo al transformismo: el impacto de Geoffroy Saint-Hilaire en la biología

Figura 1.12: Retrato de Jean Baptiste Lamarck. Imagen disponible bajo licencia Creative Commons CC0.
Geoffroy Saint-Hilaire (1772-1884) (Fig. 1.12), discípulo de Cuvier, desempeñó un papel destacado en la evolución del pensamiento científico en París durante el siglo XVIII y principios del XIX. Inicialmente, adoptó una postura fijista, siguiendo la línea de su mentor Cuvier. Sin embargo, con el tiempo, evolucionó hacia una postura transformista, influenciado por las ideas de Lamarck. En su visión transformista, Geoffroy Saint-Hilaire argumentó que las especies podían transformarse como resultado de cambios ambientales, especialmente aquellos relacionados con el enfriamiento de la Tierra. Este enfoque marcó un desvío de la posición fijista predominante en la época. Geoffroy Saint-Hilaire también investigó el desarrollo embrionario en busca de evidencias de la transformación de las especies. Su trabajo en embriología contribuyó a su perspectiva transformista y fortaleció su convicción de que las especies no eran inmutables. Geoffroy Saint-Hilaire investigó las malformaciones congénitas (teratologías) (Fig. 1.13), propuso que seguían patrones y leyes del desarrollo y creía que estas podían proporcionar información sobre la evolución y la función de las estructuras anatómicas. Su trabajo sugirió que las diferencias en la forma y función entre las especies podrían deberse a variaciones en el desarrollo embrionario y, en última instancia, influir en la evolución de esas especies. Esta perspectiva influyó en la comprensión de que las variaciones en las características de los organismos, incluidas las malformaciones, podían formar la base para la evolución de nuevas especies a lo largo del tiempo.
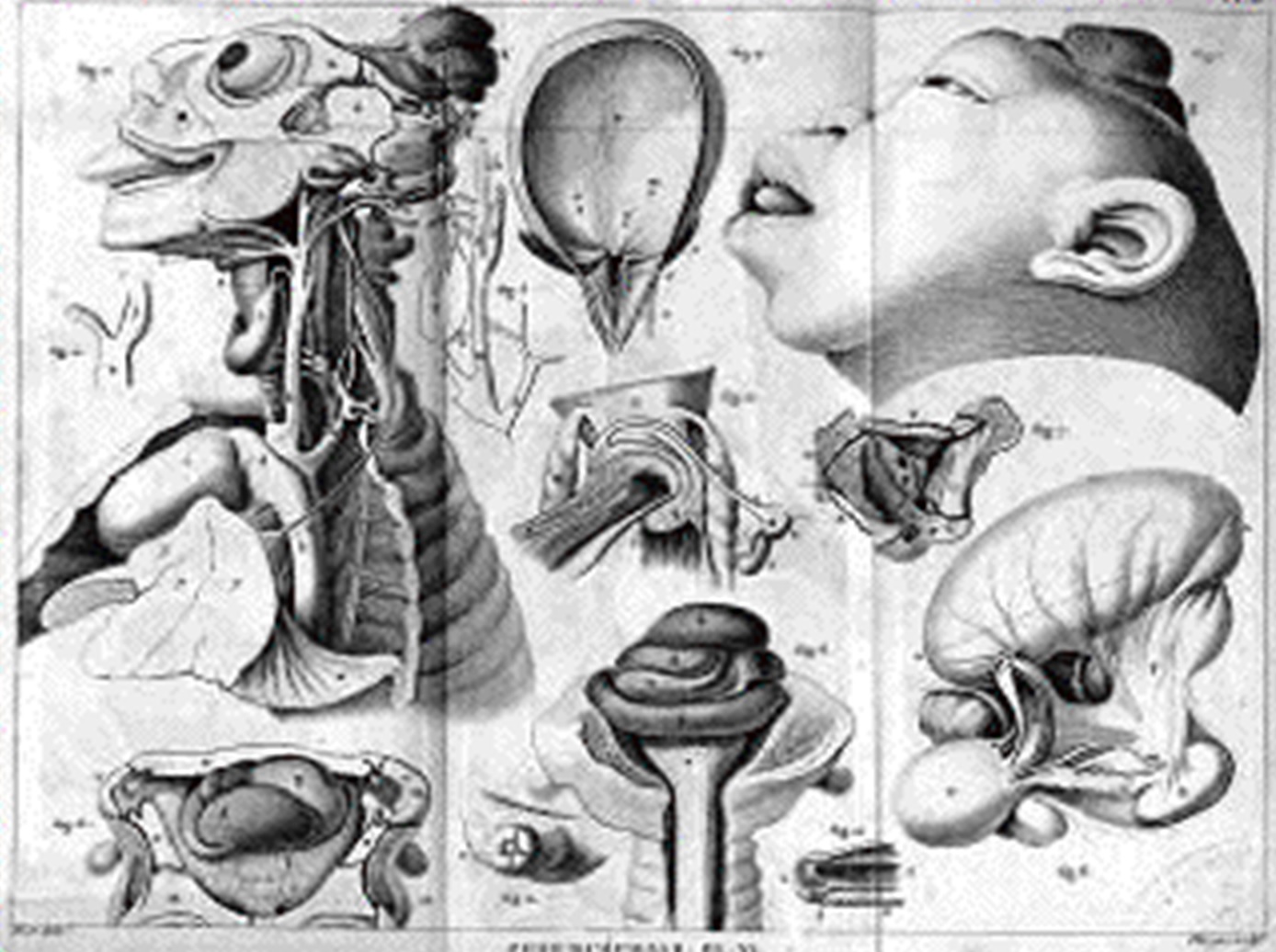
Figura 1.13: Dibujo de una teratología de Geoffroy Saint-Hilaire. Imagen disponible bajo licencia Creative Commons CC0.
La “Teoría de los Análogos” y el “Principio de las Conexiones” son conceptos clave en la obra de Geoffroy Saint-Hilaire. Estos conceptos se relacionan con la idea de que existe una unidad en el plan corporal de los animales y que las partes del cuerpo tienen conexiones anatómicas y evolutivas significativas.
Teoría de los Análogos: Esta teoría se basa en la idea de que existe una correspondencia o similitud en la estructura de los órganos entre diferentes especies, a pesar de las diferencias morfológicas externas. En otras palabras, según la teoría de los análogos, los órganos homólogos en diferentes especies, a pesar de sus adaptaciones y funciones específicas, tienen un modelo subyacente común.
Principio de las Conexiones: Este principio se refiere a la idea de que las partes del cuerpo de un organismo están relacionadas y conectadas entre sí a nivel anatómico y evolutivo. Geoffroy Saint-Hilaire argumentaba que las diferencias anatómicas en las especies se debían a la variación en la forma en que estas conexiones se desarrollaban durante el desarrollo embrionario. Esto sugiere que el cambio en una parte del cuerpo de un organismo puede influir en otras partes del cuerpo debido a estas conexiones.
1.1.16 Siglo XVIII y principios del Siglo XIX en Gran Bretaña
1.1.16.1 William Smith y la revolución geológica en Gran Bretaña: el legado del padre de la geología inglesa

Figura 1.14: Retrato de William Smith. Imagen disponible bajo licencia Creative Commons CC0.
La transición del siglo XVIII al XIX marcó un período crucial en la evolución del pensamiento científico en Gran Bretaña, particularmente en el campo de la geología. Entre las destacadas figuras de esta época, William Smith (1769-1839) (Fig. 1.14) se destacó como una figura influyente que transformó la comprensión de la geología inglesa. Conocido como el “Padre de la geología inglesa”, Smith dejó un legado perdurable a través de sus contribuciones pioneras que abrieron la puerta a una nueva era en la geología y la paleontología.
Una de las contribuciones más significativas de William Smith fue su demostración de que los fósiles son indicadores mucho más confiables de los estratos que la litología, una idea revolucionaria que cambió la forma en que se entendía la geología. Hasta ese momento, la litología, o la composición de las rocas, se consideraba el principal criterio para la clasificación de estratos geológicos. Sin embargo, Smith observó que los fósiles presentes en las capas de roca variaban de manera predecible y característica, lo que le permitió desarrollar un sistema de estratigrafía basado en la sucesión de fósiles. Esta observación condujo a la creación del primer mapa geológico de Inglaterra y Gales, que trazó la distribución de estratos y fósiles en la región (Fig. 1.15).
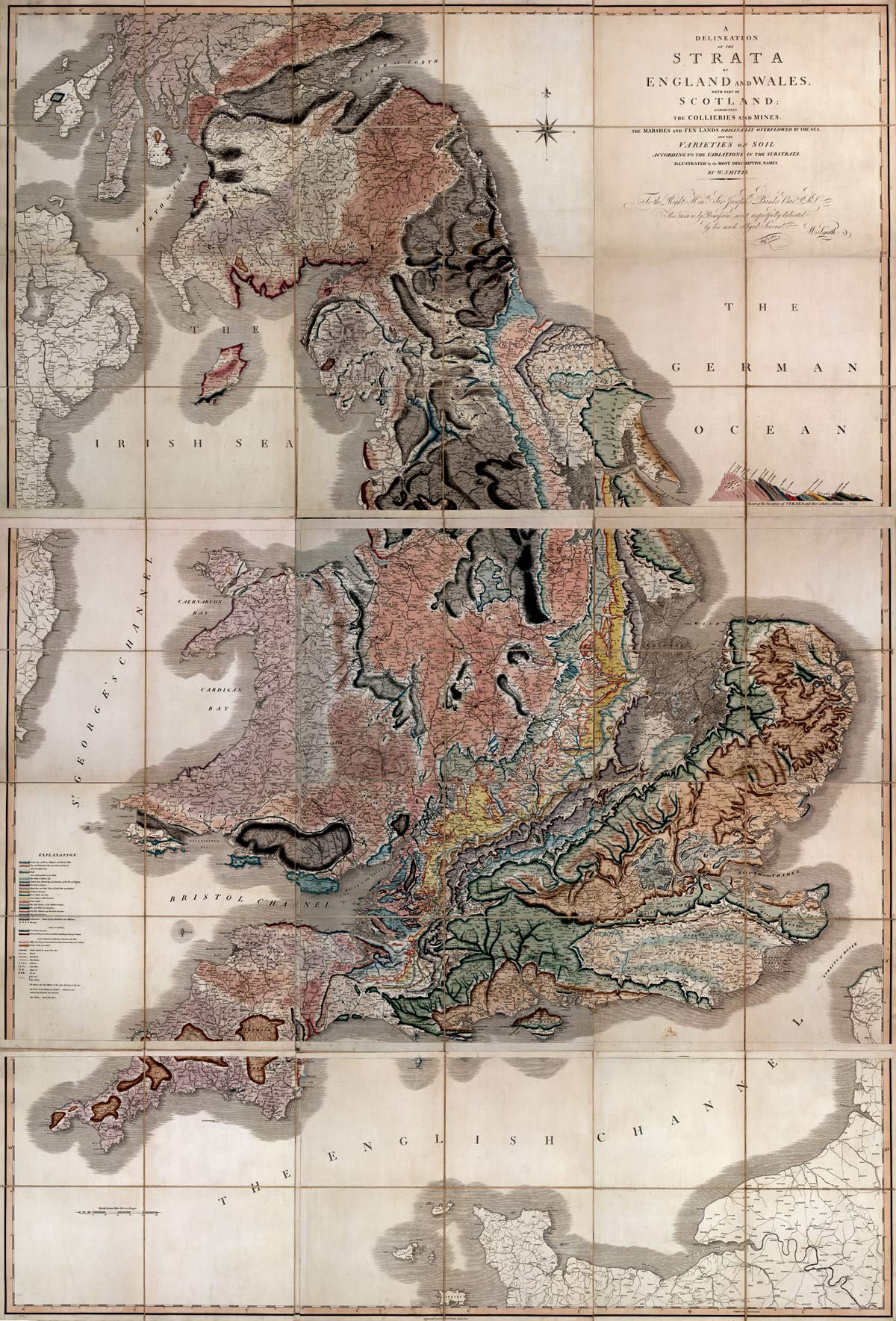
Figura 1.15: Mapa geológico de William Smith. Imagen disponible bajo licencia Creative Commons CC0.
La influencia de Smith trascendió los confines de sus mapas geológicos. Contribuyó en gran medida a la fundación de la Geological Society of London, que rápidamente se convirtió en el principal foro de debate paleontológico y geológico en lugar del Museo de París. En esta sociedad, Smith interactuó con otros notables paleontólogos y geólogos de la época, incluidas figuras como Murchison y Parkinson, enriqueciendo así el intercambio de ideas y avances científicos.
En resumen, William Smith desempeñó un papel fundamental en la revolución geológica de Gran Bretaña durante el siglo XVIII y principios del XIX. Su reconocimiento de los fósiles como marcadores confiables de los estratos cambió la forma en que se abordaba la geología, y su legado perdura en la ciencia moderna. Aunque su perspectiva a menudo se fusionaba con creencias teológicas, Smith abrió las puertas a una comprensión más profunda de la historia de la Tierra y dejó una huella indeleble en la geología inglesa.
1.1.16.2 William Buckland: la influencia de Cuvier y la teología en la geología británica del siglo XIX

Figura 1.16: Retrato de William Buckland. Imagen disponible bajo licencia Creative Commons CC0.
A pesar de los avances científicos, la influencia de Georges Cuvier seguía siendo influyente en Gran Bretaña, y la teología desempeñó un papel importante en la concepción geológica de muchos científicos británicos. William Buckland (1784-1856) (Fig. 1.16), un destacado geólogo de la época, desempeñó un papel importante en la traducción al inglés de la obra de Cuvier relacionada con la cronología bíblica. Este esfuerzo no solo contribuyó a la difusión de las ideas de Cuvier, sino que también consolidó una inclinación teológica en la comunidad geológica británica. La interacción de la geología con la teología se reflejó en la creencia en la teoría del Diluvio, que Buckland defendió como el principal defensor en Gran Bretaña. Esta teoría sostenía que un diluvio bíblico había dado forma a la Tierra y sus características geológicas, un enfoque que enfatizaba la intervención divina en la historia geológica. La perspectiva de Buckland también incluía la idea de progresionismo, que sostenía que las formas de vida imperfectas eran reemplazadas por formas mejoradas a lo largo del tiempo. Esta concepción del cambio y la mejora continuos estaba en consonancia con sus creencias religiosas y contribuyó a la interacción entre la geología y la teología.
1.1.16.3 Charles Lyell: uniformismo y su influencia en la geología y la biología

Figura 1.17: Retrato de Charles Lyell. Imagen disponible bajo licencia Creative Commons CC0.
Charles Lyell (1797-1875) (Fig. 1.17), conocido como el padre del uniformismo, desempeñó un papel fundamental en la evolución del pensamiento científico en el siglo XIX, influyendo tanto en la geología como en la biología. Lyell, discípulo de Buckland, abogó por la idea de que los procesos geológicos y naturales que observamos en la actualidad han estado en funcionamiento durante toda la historia de la Tierra, sin cambios significativos.
Lyell argumentó que no había evidencia de que la vida, el clima, los ambientes y los procesos geológicos hubieran sido diferentes en el pasado. Sostenía que no había señales de progresión en el registro fósil y negaba la idea de que los fósiles representaban formas inferiores a las actuales. Además, rechazó la necesidad de explicar eventos catastróficos como la causa de los cambios geológicos, afirmando que la ausencia de sedimentación continua podía dar cuenta de lo que parecían “catástrofes”.
Una de las contribuciones más significativas de Lyell fue su obra “Principles of Geology” (1830-1833), que ejerció una gran influencia en la comunidad científica, incluyendo a Charles Darwin. A su regreso del viaje del Beagle, Darwin entabló amistad con Lyell, quien le proporcionó acceso a la Geological Society y lo inspiró con sus ideas sobre la antigüedad de la Tierra y la constancia de los procesos geológicos. Esta influencia fue crucial para el desarrollo de la teoría de la evolución de Darwin.
Sin embargo, las ideas del uniformismo se tambalearon cuando se estudiaron materiales anteriores al Carbonífero. El Silúrico, caracterizado por una fauna cálida y la ausencia de plantas, y el Cámbrico, que presentaba organismos complejos, contradecían la noción de un ritmo de cambio uniforme. Estos descubrimientos desafiaron la creencia en un mundo geológico estático y llevaron a una reevaluación de las teorías geológicas y biológicas.
1.1.17 Elementos clave para la comprensión de la evolución en el siglo XIX
El siglo XIX marcó un período de transformación y avances significativos donde diversas figuras destacadas contribuyeron a la emergencia de los elementos clave para la comprensión evolutiva de la vida, incluyendo homologías, parecidos embrionarios y el registro fósil.
Robert Chambers (1802-1871) propuso la controvertida idea de que el origen de las especies podría no estar directamente vinculado a la intervención divina, desafiando las creencias religiosas tradicionales. Este planteamiento abrió el camino a una discusión más amplia sobre la evolución. Su libro anónimo, Vestiges of the Natural History of Creation (1844), alcanzó gran popularidad, llegando a su 12ª edición antes de que se revelara su identidad como autor. En la obra, Chambers presentaba una teoría del origen cósmico, sugiriendo que cada forma natural tenía una predecesora y defendía la generación espontánea basándose en experimentos con electricidad e insectos. Aunque el libro contenía errores científicos, influyó profundamente en figuras como Charles Darwin y Alfred Russel Wallace. Darwin reconoció que Vestiges preparó a la sociedad para aceptar hipótesis evolutivas, mientras que Wallace lo consideró una hipótesis plausible que lo impulsó a buscar evidencia sobre la evolución durante su trabajo de campo.
Heinrich Georg Bronn (1800-1862) desempeñó un papel fundamental al predisponer a los paleontólogos a comprender el registro fósil en términos de la derivación de especies a partir de otras preexistentes. Esta perspectiva ayudó a sentar las bases para la comprensión de la evolución de las especies a lo largo del tiempo.

Figura 1.18: Retrato de Richard Owen. Imagen disponible bajo licencia Creative Commons CC0.
Richard Owen (1804-1892) (Fig. 1.18) formuló la noción de homología y arquetipo o plan corporal, influenciado por las ideas de Saint-Hilaire. La idea de homología, que sugiere similitudes anatómicas entre diferentes especies debido a un ancestro común, fue fundamental para la comprensión de la evolución de las estructuras biológicas. La noción del arquetipo o plan corporal destacó la presencia de una estructura básica compartida entre las especies (Fig. 1.19).
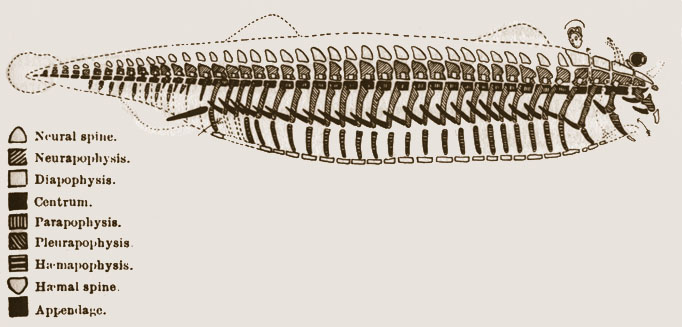
Figura 1.19: Arquetipo de vertebrado según Richard Owen. Imagen disponible bajo licencia Creative Commons CC0.
La obra de Owen contribuyó a consolidar estos conceptos en la biología evolutiva, aunque las correspondencias entre órganos y estructuras se conocían de mucho antes. Por ejemplo, destacan las contribuciones de Pierre Belon en el siglo XVI. En su obra “La historia de la naturaleza de las aves” (1555), incluyó dos figuras de los esqueletos de humanos y aves, señalando los huesos homólogos. Esto se considera ampliamente como una de las primeras demostraciones de anatomía comparada (Fig. 1.20).
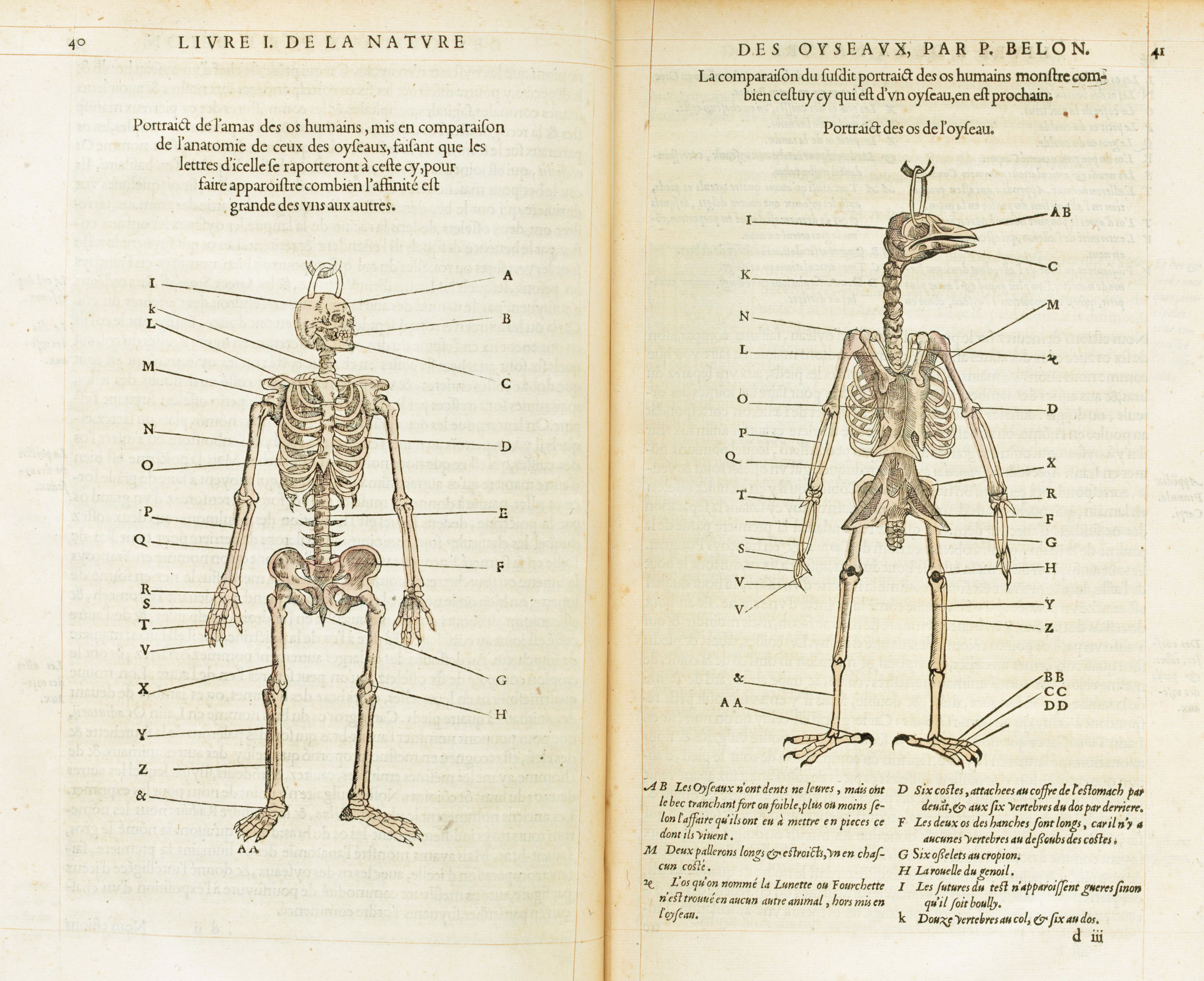
Figura 1.20: Comparativa entre esqueletos de un humano y un ave de Pierre Belon. Imagen disponible bajo licencia Creative Commons CC0.
1.1.17.1 Charles Darwin: selección satural, uniformidad y registro fósil en el siglo XIX

Figura 1.21: Retrato de Charles Darwin. Imagen disponible bajo licencia Creative Commons CC0.
En el siglo XIX, la figura de Charles Darwin (1809-1882) (Fig. 1.21) y su viaje a bordo del HMS Beagle entre 1831 y 1836 marcaron un hito en la historia de la biología. Durante este viaje, Darwin recopiló una cantidad impresionante de datos biogeográficos, ecológicos y sistemáticos que se convertirían en la base de sus revolucionarias ideas sobre la evolución.
Uno de los aspectos clave de las observaciones de Darwin fue su enfoque en la geología, guiado por el concepto de la uniformidad propuesto por Charles Lyell que le proporcionó la base conceptual necesaria para comprender cómo las especies podrían haber evolucionado gradualmente a lo largo del tiempo. Al aplicar la uniformidad a la biología, Darwin sugirió que, al igual que los procesos geológicos que actúan lentamente pueden dar forma a la Tierra, los procesos biológicos a lo largo de generaciones pueden dar forma a la vida en la Tierra.
Darwin no se conformó con explicaciones superficiales o teológicas, como las propuestas por Robert Chambers. En cambio, se embarcó en el desafío de responder a dos de las preguntas más fundamentales de la biología: “¿por qué” y “cómo” algunas especies se derivan en otras por modificación. Refinó sus argumentos para abordar estas cuestiones de manera más profunda y científica, alejándose de las explicaciones superficiales que no proporcionaban una comprensión completa del proceso evolutivo. En su obra icónica “On the origin of species” (Fig. 1.22), publicada en 1859, Charles Darwin presentó una serie de argumentos fundamentales que sustentaban su teoría de la evolución.

Figura 1.22: Portada de la edición original de 1859 de On the origin of species de Charles Darwin. Imagen disponible bajo licencia Creative Commons CC0.
Uno de los argumentos centrales de Darwin se basó en la observación de la selección artificial, una práctica conocida por agricultores y ganaderos. Darwin notó que los seres humanos habían estado realizando selección artificial durante siglos, eligiendo y criando deliberadamente plantas y animales con ciertas características deseables. Al aplicar este concepto a la naturaleza, Darwin razonó que si los seres humanos podían influir en la evolución de las especies mediante la selección artificial, entonces la naturaleza también podía ejercer una forma de selección, aunque de manera no intencionada. Esta idea sentó las bases para su concepto de “selección natural,” donde las especies evolucionan a lo largo del tiempo debido a la presión de selección ejercida por el entorno y la competencia por los recursos limitados.
El argumento de la limitación de los recursos y su influencia en la demografía humana, que estuvo influenciado por las ideas del economista Thomas Malthus, fue otro pilar importante de la teoría de Darwin. Malthus sostenía que, en un mundo con recursos limitados, la población humana tiende a crecer más rápido que la capacidad de la Tierra para proporcionar alimentos y otros recursos. Darwin aplicó este principio a la naturaleza y argumentó que en un mundo con recursos limitados, la naturaleza selecciona a los organismos que son más capaces, debido a sus características, de utilizar esos recursos de manera ventajosa. Estas características, que proporcionan una ventaja en la lucha por la supervivencia y la reproducción, se transmiten de generación en generación, ya que son heredables. Esto llevó a la idea central de la “selección natural” de Darwin: las características beneficiosas se acumulan en las poblaciones a lo largo del tiempo, lo que conduce a la evolución de las especies.
En la época de Charles Darwin, las ideas sobre la uniformidad de la naturaleza y el registro fósil jugaron un papel crucial en su desarrollo de la teoría de la evolución. Darwin se encontró con un enigma en la naturaleza que trataba de resolver a través de su comprensión de la uniformidad de los procesos naturales y la evidencia del registro fósil. La “naturaleza uniforme” implicaba que los procesos geológicos y biológicos en la Tierra ocurrían de manera constante y a un ritmo constante a lo largo del tiempo. Esto requería que cualquier transformación de las especies ocurriera en pasos imperceptibles y a una velocidad muy lenta. Darwin llegó a comprender que los procesos de selección natural, que actuaban gradualmente a lo largo de generaciones sucesivas, eran consistentes con esta idea de uniformidad. En otras palabras, los cambios en las especies serían tan sutiles que serían difíciles de detectar en una escala de tiempo humana.
Sin embargo, Darwin enfrentó un desafío importante al considerar el registro fósil. A primera vista, el registro fósil parecía contradecir su teoría. No se encontraban fósiles que documentaran las numerosas formas de transición gradual que él esperaba ver si la evolución era un proceso constante y gradual. En lugar de eso, parecía que se presentaban períodos de aparición súbita de nuevas formas seguidos por largos períodos de estasis, donde las especies parecían mantenerse sin cambios durante millones de años. Esto llevó a Darwin a ver el registro fósil como una evidencia en contra de sus ideas en un principio. Además, en la época de Darwin, se estimaba que la Tierra tenía una edad de alrededor de 98 millones de años, lo que parecía incompatible con las tasas de cambio que se requerirían para explicar la diversidad de la vida a través de la selección natural a lo largo del tiempo.
1.1.17.1.1 Resumen de los capítulos de “El Origen de las Especies”
La variación en estado doméstico. Darwin empieza describiendo cómo las especies varían bajo domesticación, como ocurre con las razas de perros, palomas o plantas cultivadas. Argumenta que los humanos han seleccionado ciertas características útiles o deseables en animales y plantas, un proceso llamado “selección artificial”. Esto le sirve como analogía para explicar cómo podría funcionar la selección natural en la naturaleza.
La variación en la naturaleza. Este capítulo explora cómo las especies varían en la naturaleza, no solo en estado doméstico. Darwin observa que existe una gran cantidad de variación en las características de los organismos, incluso dentro de la misma especie. Estas variaciones son esenciales para la selección natural, ya que las diferencias entre los individuos pueden hacer que algunos sobrevivan mejor que otros.
La lucha por la existencia. Darwin introduce el concepto de la “lucha por la existencia”, influido por la lectura de Thomas Malthus. Dado que las especies tienden a producir más descendencia de la que puede sobrevivir, existe una competencia por los recursos limitados, lo que lleva a que solo los organismos mejor adaptados sobrevivan y se reproduzcan. Aquí empieza a desarrollar la idea de la selección natural.
La selección natural. Este capítulo es central en su obra. Darwin explica cómo la selección natural actúa sobre la variación en las especies, favoreciendo a los individuos que poseen características que los hacen más aptos para su entorno. Esos individuos tienen más probabilidades de sobrevivir y reproducirse, pasando sus características ventajosas a las siguientes generaciones. Así, la selección natural es el mecanismo por el cual las especies evolucionan.
Leyes de variación. Darwin explora las causas de la variación en los organismos, aunque admite que no se conoce completamente en su tiempo. Señala que la variación puede depender de varios factores, incluidos el ambiente, la herencia, el uso y el desuso de partes del cuerpo, y las condiciones de vida. También introduce la noción de “correlación de crecimiento”, sugiriendo que un cambio en una parte del organismo puede influir en otras partes.
Dificultades de la teoría. Darwin aborda las posibles objeciones a su teoría. Estas incluyen preguntas como: ¿Cómo podrían evolucionar estructuras complejas como el ojo? ¿Cómo se explican las variedades intermedias si la selección natural actúa gradualmente? Darwin reconoce las dificultades pero ofrece explicaciones basadas en la gradualidad de la evolución y la acumulación de cambios pequeños.
Objeciones diversas a la teoría de la selección natural. Este capítulo responde a otras críticas, como la falta de variedades intermedias y la estabilidad de las especies. Darwin admite que la falta de fósiles intermedios es un problema, pero sugiere que el registro fósil es incompleto y que muchas formas transitorias no se han preservado.
Instinto. Darwin compara los instintos de los animales con los hábitos adquiridos por el hombre y analiza cómo los comportamientos complejos, como el de las abejas haciendo panales, pudieron haber evolucionado a través de la selección natural. Sostiene que los instintos son análogos a las estructuras corporales y pueden evolucionar de manera gradual.
Hibridismo. Aquí Darwin estudia la fertilidad de los híbridos y cómo la hibridación afecta a la evolución. Reconoce que la hibridación puede ser problemática para la selección natural, pero explica que, en la mayoría de los casos, los híbridos entre especies diferentes suelen ser estériles o menos viables, lo que ayuda a mantener las especies separadas.
Sobre la imperfección del registro geológico. Darwin explica que el registro fósil es incompleto, lo que puede explicar la falta de formas intermedias entre especies en el registro geológico. También describe cómo las capas geológicas y los procesos de fosilización afectan la preservación de organismos antiguos, y sugiere que muchas formas transitorias no se han conservado.
Sobre la sucesión geológica de los seres orgánicos. Este capítulo analiza cómo las especies cambian a lo largo del tiempo en el registro fósil. Darwin observa que las especies extintas están relacionadas con las formas vivas actuales y que los cambios evolutivos ocurren gradualmente, en lugar de a través de eventos catastróficos, como se pensaba anteriormente.
Distribución geográfica. Darwin investiga la distribución de las especies en todo el mundo. Señala que las especies que viven en islas aisladas son a menudo muy diferentes de las del continente más cercano, lo que apoya su teoría de la evolución. También sugiere que los patrones de migración y los cambios geológicos han afectado la dispersión de las especies. XII
Afinidades mutuas de los seres orgánicos: morfología, embriología, órganos rudimentarios. Este capítulo explora las similitudes entre los seres vivos. Darwin estudia la morfología (forma) y la embriología (desarrollo temprano) de los organismos, mostrando que los organismos que están relacionados evolutivamente comparten patrones similares de desarrollo y anatomía. También menciona los “órganos rudimentarios” o vestigiales (como los restos de patas en serpientes), que son vestigios de un pasado evolutivo.
Recapitulación y conclusión. Darwin recapitula sus ideas sobre la selección natural y la evolución. Afirma que la evolución por selección natural es la mejor explicación para la diversidad de la vida en la Tierra. Insiste en que la selección natural es un proceso lento, acumulativo y gradual, que explica cómo las especies cambian con el tiempo, adaptándose a su entorno. Finaliza sugiriendo que su teoría puede aplicarse a todas las formas de vida y que está abierta a ser revisada a medida que se descubran más pruebas.
1.1.17.2 Alfred Russel Wallace: el descubrimiento independiente de la selección natural

Figura 1.23: Portada de la edición original de 1859 de On the origin of species de Charles Darwin. Imagen disponible bajo licencia Creative Commons CC0.
En el siglo XIX, la figura de Alfred Russel Wallace (1823-1913) (Fig. 1.23), un naturalista de origen galés, desempeñó un papel fundamental en el desarrollo de la teoría de la evolución, aunque a menudo su contribución no recibe el reconocimiento que merece. Su historia se entrelaza de manera notable con la de Charles Darwin, ya que ambos, de manera independiente, concebieron el principio de la selección natural, un mecanismo crucial en la teoría de la evolución.
La historia comienza con Charles Darwin, quien comenzó a desarrollar su teoría de la selección natural a partir de 1838. Durante décadas, Darwin trabajó arduamente en su “gran libro” sobre la evolución, que se convertiría en su obra maestra, “El Origen de las Especies”. Mientras tanto, en una parte distante del mundo, Alfred Russel Wallace estaba llevando a cabo investigaciones en el sudeste asiático, recolectando datos y observaciones sobre la diversidad de la vida en la región.
En un momento crítico, Wallace llegó de manera independiente a una conclusión sorprendentemente similar a la de Darwin: las especies evolucionan a lo largo del tiempo debido a la selección natural. Wallace resumió sus ideas en un ensayo y se lo envió a Darwin para obtener su opinión. Darwin se sorprendió al recibir este trabajo, ya que sus investigaciones paralelas habían llegado a una conclusión casi idéntica. La respuesta de Darwin ante esta coincidencia es notable y ejemplar. En lugar de competir con Wallace, Darwin decidió presentar ambas teorías en conjunto. En 1858, los trabajos de Darwin y Wallace se presentaron en una reunión conjunta de la Sociedad Linneana de Londres. Aunque Darwin ya había formulado sus ideas sobre la selección natural en años anteriores, fue reconocido como el primero en presentar estas ideas por Lyell y Hooker, dos prominentes científicos de la época. Utilizaron como prueba un ensayo de Darwin de 1844 y una carta que había enviado a Asa Gray en 1857, ambos publicados junto con el artículo de Wallace.
Este evento fue un hito en la historia de la biología. La selección natural se convirtió en un componente crucial para entender la evolución y Darwin y Wallace compartieron el reconocimiento por su formulación independiente de este mecanismo. Es en este contexto que se ha propuesto la frase “el mecanismo de Darwin/Wallace de la selección natural” para enfatizar la importancia de ambos científicos en el desarrollo de esta teoría.
1.1.17.3 Al-Jahiz: raíces históricas de la selección natural

Figura 1.24: Retrato de Al-Jahiz. Imagen disponible bajo licencia Creative Commons CC0.
Es relevante destacar que, si bien Darwin y Wallace son los nombres más conocidos en relación con la teoría de la selección natural en el siglo XIX, ideas similares sobre la evolución habían sido postuladas con anterioridad. Por ejemplo, en el siglo IX, el pensador islámico Al-Jahiz presentó una argumentación similar en su obra “El libro de los animales”, lo que demuestra que el concepto de la selección natural tenía raíces en la reflexión humana mucho antes de que se desarrollara como parte de una teoría científica bien fundamentada.
Al-Jahiz, cuyo nombre completo es Abu Uthman Amr ibn Bahr al-Kinani al-Fuqaimi al-Basri, fue un influyente escritor, filósofo, y erudito árabe que vivió durante el siglo IX (776-869) (Fig. 1.24). Es especialmente conocido por su obra “Kitab al-Hayawan” (El Libro de los Animales). Al-Jahiz nació en Basora, en la actual Irak, y su obra abarcó una variedad de temas, incluyendo biología, zoología, lingüística, retórica, teología y filosofía.
“El Libro de los Animales” (Fig. 1.25), la obra más destacada de Al-Jahiz, es un extenso tratado de zoología que aborda una amplia variedad de temas relacionados con el mundo animal. En este trabajo, Al-Jahiz observó y analizó el comportamiento y las características de numerosas especies, describiendo en detalle sus atributos físicos y sus conductas. Aunque su enfoque fue principalmente descriptivo, también exploró conceptos relacionados con la evolución. Formuló ideas pioneras sobre la lucha por la existencia y la adaptación en el reino animal, aunque no desarrolló un marco teórico coherente y estructurado sobre la evolución, como lo hicieron Charles Darwin y Alfred Russel Wallace siglos más tarde.
En “El Libro de los Animales”, Al-Jahiz defendió la idea de que las especies de animales cambiaban con el tiempo en respuesta a las presiones ambientales y que solo los animales mejor adaptados sobrevivían y se reproducían. Sus observaciones sobre la competencia y la adaptación entre las especies prefiguraron conceptos clave de la teoría de la evolución.

Figura 1.25: Paginas de El Libro de los Animales de Al-Jahiz. Imagen disponible bajo licencia Creative Commons CC0.
Además de sus contribuciones a la zoología y la biología, Al-Jahiz también fue un prolífico escritor de ensayos, libros de viajes y obras sobre retórica, lingüística y teología. Su trabajo en la lengua árabe y la literatura ayudó a establecer fundamentos importantes para el estudio de la lengua y la cultura árabe.
Lectura recomendada
Álvarez, F. (2007). El Libro de los Animales de al-Jahiz, un esbozo evolucionista del siglo IX Evolución, 2, 25-29.
1.1.18 La recepción de la teoría de la evolución por parte de los paleontólogos del siglo XIX
En el siglo XIX, la idea de la evolución provocó una serie de reacciones en la comunidad de paleontólogos, que estaban en proceso de descubrir y comprender la diversidad de formas de vida en la Tierra a través del registro fósil. A medida que la teoría de la evolución de Charles Darwin comenzó a tomar forma y a difundirse, algunos paleontólogos se sintieron intrigados y alentados por la posibilidad de encontrar evidencia de esta teoría en los fósiles.

Figura 1.26: Retrato de Thomas Huxley. Imagen disponible bajo licencia Creative Commons CC0.
Uno de los personajes destacados de esta época fue Thomas Henry Huxley (1825-1895) (Fig. 1.26), un biólogo y filósofo británico conocido como el “Bulldog de Darwin” debido a su firme defensa de la teoría de la evolución de Charles Darwin. Huxley alentó a Darwin a buscar formas transicionales en el registro fósil, especialmente por encima del nivel de especie. Esta sugerencia impulsó a Darwin a explorar aún más la evidencia fósil en busca de pruebas que respaldaran su teoría.
Uno de los hallazgos paleontológicos más notables de esta época fue el descubrimiento del Archaeopteryx en Solnhofen, Alemania (Fig. 1.27). Este fósil, datado en el Jurásico tardío, presentaba características tanto de dinosaurio como de ave, lo que lo convirtió en un ejemplo claro de una forma transicional entre estos dos grupos de organismos proporcionando una evidencia convincente de la evolución de las aves a partir de dinosaurios.

Figura 1.27: Especimen de Archaeopteryx lithographica del Museum für Naturkunde de Berlín. Imagen disponible bajo licencia Creative Commons CC0.
En el ámbito de la paleobiogeografía, Albert Gaudry (1827-1908) hizo un descubrimiento relevante al encontrar fósiles de Hipparion en el Mioceno superior. Hipparion era una forma transicional entre los équidos del Eoceno de París y los pleistocenos, lo que sugiere una evolución en la línea de los caballos a lo largo del tiempo. Este hallazgo representó una de las primeras interpretaciones evolutivas basadas en evidencia fósil y apoyó firmemente la idea de que las especies podían cambiar con el tiempo.
Otro paleontólogo destacado, Othniel Charles Marsh (1831-1899), llevó a cabo estudios exhaustivos de équidos americanos y encontró una multitud de formas intermedias en el registro fósil. Además, propuso hipótesis paleobiogeográficas para explicar la distribución geográfica de estas especies. Sus investigaciones proporcionaron más evidencia de la evolución y cómo las especies podrían cambiar gradualmente a lo largo del tiempo y el espacio.
Otro de los desafíos más apremiantes en el campo de la paleontología y la biología fue comprender y documentar las transiciones tanto entre especies como entre filos en el registro fósil. En esa época, muchos científicos creían que el registro fósil era tan imperfecto, debido a la preservación limitada de fósiles y a su naturaleza fragmentaria, que sería imposible encontrar evidencia de formas transicionales entre especies o “antepasados” de grandes grupos. Sin embargo, varios descubrimientos sorprendentes rompieron con esa perspectiva y ayudaron a respaldar la idea emergente de la evolución.
Uno de los primeros y más claros ejemplos de evolución gradual a nivel de especies documentada es el trabajo del paleontólogo austrohúngaro Melchior Neumayr (1845–1890), quien en sus estudios de moluscos de agua dulce del Terciario superior de la actual Europa del Este mostró la transformación gradual de la morfología de las conchas a través de diferentes estratos (Fig. 1.28). Neumayr, seguidor de Darwin, fue pionero en demostrar cómo la variación y evolución de las especies podían rastrearse en fósiles de invertebrados, proporcionando una evidencia clara del proceso de evolución gradual.

Figura 1.28: Lamina ilustrando moluscos de agua dulce del trabajo de Melchior Neumayr. Imagen disponible bajo licencia Creative Commons CC0.
Otro ejemplo significativo de evolución gradual a nivel de especie es el estudio de Micraster realizado por el paleontólogo británico Arthur Rowe (1858–1926) en 1899 (Fig. 1.29). Rowe analizó alrededor de 2000 especímenes de este género a partir de depósitos de caliza del Reino Unido, documentando variaciones morfológicas a lo largo de una sucesión estratigráfica. Su enfoque innovador, que incluía descripciones cuantitativas y fotografías de alta calidad, estableció un referente en el estudio de la evolución de los equinodermos. Sin embargo, investigaciones posteriores revelaron que los patrones evolutivos en Micraster eran más complejos, sugiriendo que la evolución no siempre sigue un camino lineal.
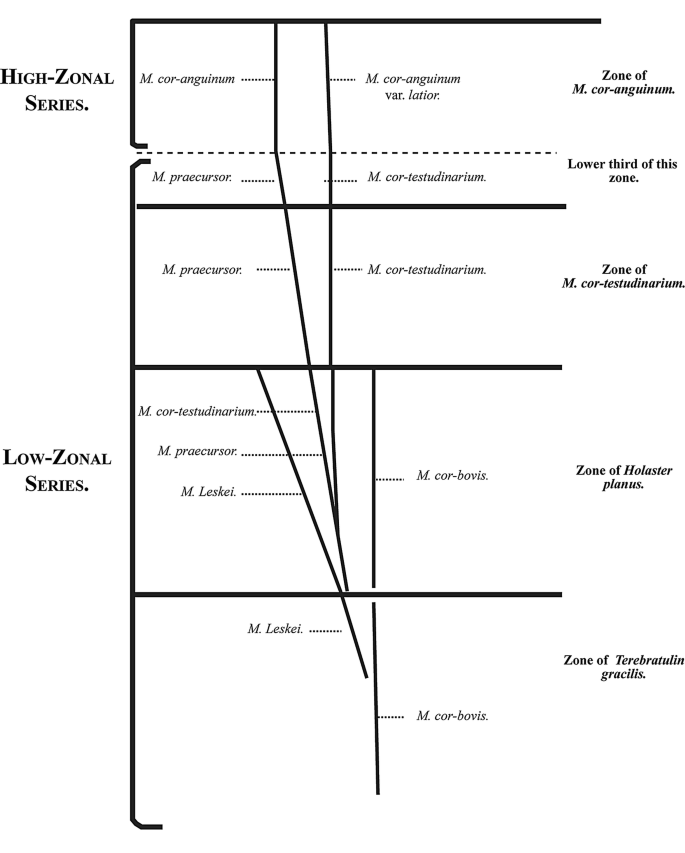
Figura 1.29: Lámina ilustrando relaciones de descendecia entre equinoideos en el trabajo de Arthur Rowe. Imagen tomada de Allmon (2020).
Además de las transiciones entre especies, también hubo un interés creciente en las transiciones entre filos en el registro fósil. En ese momento, se pensaba que el metamorfismo de los estratos precámbricos había eliminado cualquier rastro de posibles antepasados de los principales filos animales. Además, tanto la anatomía comparada como la embriología no proporcionaban evidencia convincente de transiciones entre los principales grupos de organismos. Sin embargo, los descubrimientos paleontológicos comenzaron a desafiar esta noción. Los estudios de fósiles en estratos precámbricos comenzaron a revelar organismos que podrían representar antepasados de los principales filos. Si bien estos hallazgos aún estaban sujetos a debate y análisis, sugirieron la posibilidad de que los antepasados de los filos animales no se habían perdido en la historia geológica.
Lectura recomendada
Allmon, W. D. (2020). Invertebrate paleontology and evolutionary thinking in the US and Britain, 1860–1940. Journal of the History of Biology, 53(3), 423-450.
Olson, E. C. (1981). The problem of missing links: today and yesterday. The Quarterly Review of Biology, 56(4), 405-442.
1.1.19 Piedras contra el tejado de la teoría de la evolución en el siglo XIX
A pesar de los avances notables en la teoría de la evolución a lo largo del siglo XIX, el camino hacia su aceptación generalizada estuvo marcado por desafíos y retrocesos que pusieron en duda su validez.
1.1.19.1 La primera piedra: aristogénesis y las visiones de Henry Fairfield Osborn

Figura 1.30: Retrato de Henry Osborn. Imagen disponible bajo licencia Creative Commons CC0.
Uno de los desafíos iniciales se centró en la percepción de la evolución como un proceso direccionado hacia la mejora, una noción internalista que postulaba que la evolución era independiente del entorno. Uno de los proponentes de esta idea fue Henry Fairfield Osborn (1857–1935) (Fig. 1.30), quien desarrolló una hipótesis llamada “aristogénesis” basada en un enfoque fisicoquímico de la evolución.
Osborn, un renombrado paleontólogo, desempeñó un papel destacado al describir al icónico Tyrannosaurus rex en 1905. Sin embargo, más allá de sus contribuciones paleontológicas, también promovió una perspectiva única sobre la evolución, que llamó “aristogénesis”. En este contexto, Osborn postuló la existencia de aristogenes, entidades fisicoquímicas en el genoplasma, que inducirían novedades evolutivas. Creía que estos aristogenes operaban como biomecanismos en el geneplasma de los organismos y que, en última instancia, eran responsables de la creación de novedades evolutivas. En contraste con las ideas darwinianas de mutación y selección natural, Osborn argumentó que los aristogenes, y no estos mecanismos, eran la fuente de la novedad evolutiva (Fig. 1.30).
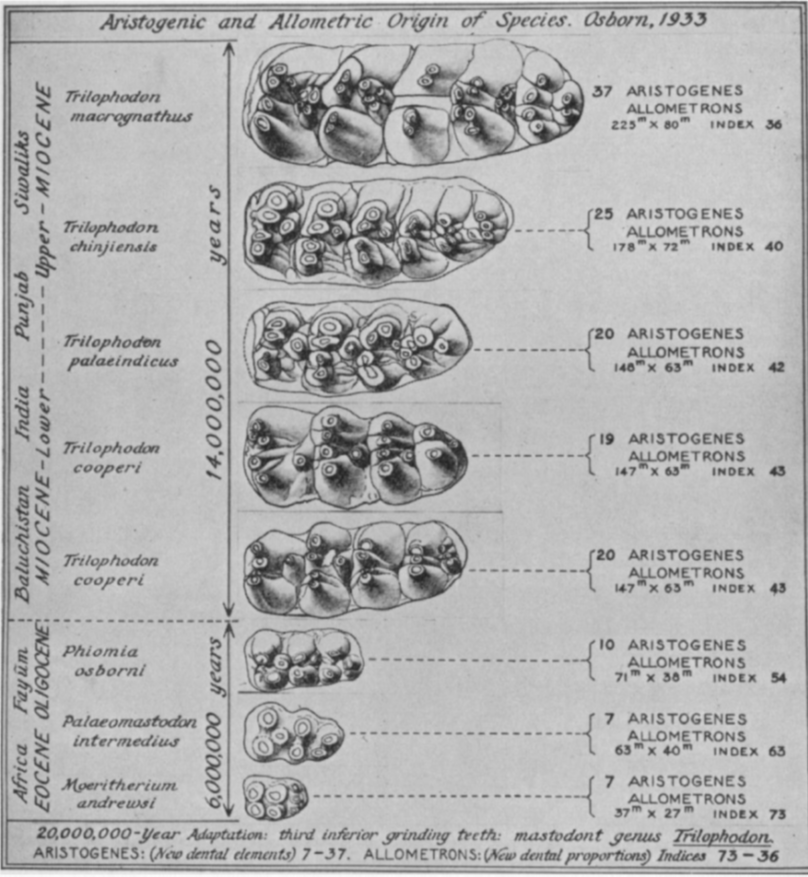
Figura 1.31: Ejemplo de aristogénesis en mastodontes según Henry Osborn. Imagen tomada de Osborn (1933).
Además de su aristogénesis, Osborn postuló su propia hipótesis sobre los orígenes humanos, conocida como la “Teoría del Hombre del Amanecer”. Basándose en el descubrimiento del Hombre de Piltdown, que luego resultó ser un engaño, Osborn propuso que los simios y los humanos compartían un ancestro común, que él creía que se había desarrollado independientemente de los simios hace millones de años. Esta idea sugiere que tanto los simios como los humanos evolucionaron en paralelo desde un ancestro común, pero Osborn enfatizó que este ancestro era más humano que simio. Su visión provocó malentendidos entre algunos, quienes creyeron erróneamente que estaba afirmando que la humanidad no había evolucionado de formas de vida inferiores.
Osborn fue inicialmente defensor del neolamarckismo propuesto por Edward Drinker Cope (padre de la famosa “regla de Cope”). Este enfoque sugería que las características adquiridas durante la vida podían heredarse, lo que implicaba un mecanismo alternativo a la selección natural darwiniana para explicar la evolución. Osborn respaldaba esta visión, que otorgaba un papel central a la adaptación individual y a la idea de un progreso biológico continuo en la evolución. Sin embargo, con el tiempo, Osborn modificó su postura y adoptó el concepto de selección orgánica, también conocido como el efecto Baldwin. Este mecanismo, propuesto por James Baldwin, sugiere que los comportamientos aprendidos por los organismos pueden influir en su éxito evolutivo, acelerando su adaptación sin que necesariamente sean heredados genéticamente en un primer momento. Más tarde, estos comportamientos podrían volverse innatos si son ventajosos, lo que integra elementos tanto darwinistas como neolamarckistas. A diferencia del neolamarckismo, que se centraba en la herencia directa de características adquiridas, la selección orgánica ofrecía una visión más integrada, donde la plasticidad fenotípica y la capacidad de aprender tenían un papel evolutivo crucial, sin violar las reglas de la herencia genética darwiniana. Una postura lamarkiana implicaría que el aprendizaje se codifica en el material genético, mientras que una darwinista implicaría que el aprendizaje, a priori, no se codifica en el material genético. El efecto Baldwin relaja esta dicotomía ya que implicaría que el aprendizaje guía la evolución por medio del proceso de asimilación genética, donde una respuesta fenotípica a un factor ambiental es asimilada (a través de la selección natural) por el genotipo de modo que se vuelve independiente del inductor ambiental.
Osborn no solo dejó su huella en la biología, sino que también desempeñó un papel influyente en las ideologías raciales de su época. En 1922, fue uno de los cofundadores de la Sociedad Estadounidense de Eugenesia, una organización que reflejaba la creencia predominante de que la herencia genética era más importante que los factores ambientales. Osborn defendía la idea de que existían razas con rasgos hereditarios inmutables y promovía la supremacía de la “raza” nórdica o anglosajona. Estas opiniones apoyaban el movimiento eugenésico, que proponía mejorar la población humana mediante la selección selectiva de individuos. Debido a su alineación con estas ideas, Osborn respaldó la obra “The Passing of the Great Race” de Madison Grant, contribuyendo con los prefacios del segundo y cuarto ediciones del libro. Esta obra, que abogaba por un sistema de selección racial basado en la eliminación de los “débiles” o “no aptos”, influyó profundamente en Adolf Hitler, quien llegó a considerarla “su biblia”.
A pesar de sus contribuciones a la paleontología y sus controvertidas opiniones sobre la eugenesia y la evolución, Henry Fairfield Osborn dejó una huella significativa en la ciencia y el pensamiento de su época, reflejando la diversidad de perspectivas que influyeron en la evolución de la teoría de la evolución en el siglo XIX.
Lectura recomendada
Bolaños Lepere, U. (2003). Simulación computacional del efecto Baldwin en un modelo depredador-presa-artificial. Instituto tecnológico y de estudios superiores de Monterrey.
Faria, F. (2017). The neo-Lamarckism of Edward Drinker Cope and the idea of biological progress in the evolutionary process. Historia Ciencias Saude-Manguinhos, 24(4), 1009-1029.
Longa, V. M. (2009). Sobre el efecto Baldwin y la noción de herencia. Signos filosóficos, 11(21), 43-72.
Osborn, H. F. (1933). Aristogenesis, the observed order of biomechanical evolution. Proceedings of the National Academy of Sciences, 19(7), 699-703.
1.1.19.2 La segunda piedra: la recapitulación filogenética de Ernst Haeckel

Figura 1.32: Ernst Haeckel en su Instituto Zoológico en Jena (Alemania). Imagen tomada de Watts et al. (2019).
En medio de las discusiones sobre la evolución en el siglo XIX, Ernst Haeckel (1834–1919)(Fig. 1.32), junto a Thomas Henry Huxley, se destacó como uno de los más fervientes defensores de las ideas de Darwin. Haeckel no solo jugó un papel crucial en la popularización del darwinismo, sino que también desarrolló la ley biogenética, afirmando que la ontogenia recapitula la filogenia: el desarrollo embrionario de un organismo repite las etapas evolutivas de su especie (Fig. 1.33). Haeckel también introdujo la teoría de la gastraea, donde postulaba que todos los animales multicelulares compartían un ancestro común, el hipotético Gastraea, cuyo desarrollo podía verse reflejado en la etapa de gastrulación en los embriones de diferentes filos animales. Estas teorías intentaban proporcionar evidencia a favor del origen monofilético de los metazoos y de la ley de la descendencia común. A través de su trabajo, Haeckel contribuyó significativamente al avance de las ciencias biológicas, especialmente en la embriología filogenética. A pesar de sus importantes contribuciones, Haeckel no estuvo exento de críticas. Sus dibujos, aunque destinados a ser ilustraciones pedagógicas más que representaciones anatómicas exactas, fueron acusados de estar estilizados o exagerados. Estas críticas se centraron en el hecho de que Haeckel a veces omitía detalles o simplificaba características embrionarias para enfatizar las similitudes entre diferentes especies. Esta simplificación fue vista por algunos como un intento de distorsionar la verdad científica, lo que generó acusaciones de fraude. Sin embargo, estas críticas han sido revaluadas en años recientes, y muchos estudiosos han defendido el valor educativo de sus ilustraciones, reconociendo que su objetivo principal era hacer que las ideas evolucionistas fueran comprensibles para un público más amplio.
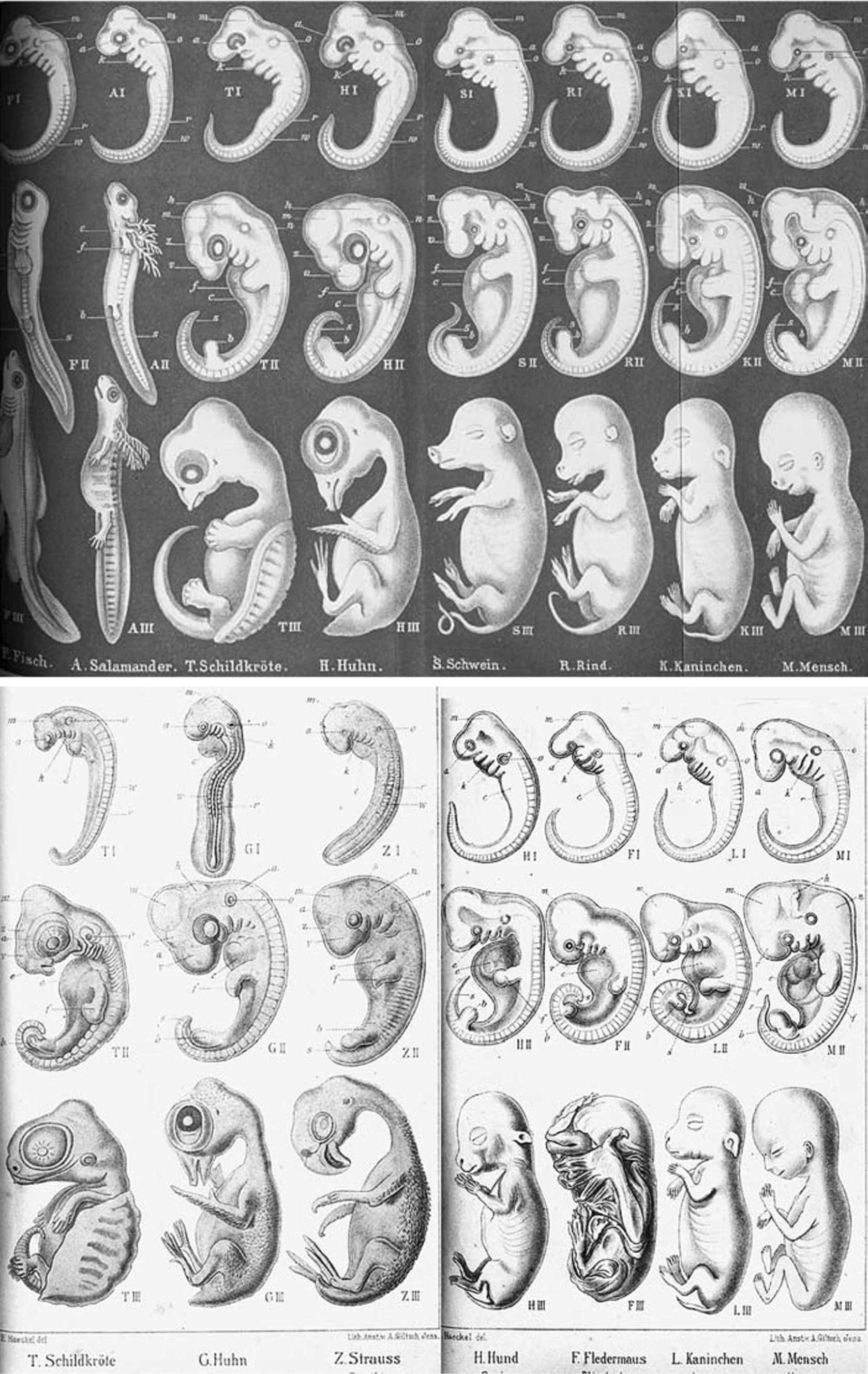
Figura 1.33: Comparación de las ilustraciones de Haeckel publicadas en 1874 (arriba) y 1891 (abajo). Imagen tomada de Watts et al. (2019).
Sumado a sus contribuciones científicas, Haeckel también fue un habilidoso artista (Fig. 1.34). Sus ilustraciones no solo sirvieron para visualizar conceptos científicos complejos, sino que también acercaron la ciencia al público general. Esta combinación de ciencia y arte hizo que sus trabajos fueran accesibles y comprensibles para audiencias más amplias, lo que fortaleció su impacto. Uno de sus mayores legados fue su capacidad para visualizar y enseñar los principios evolutivos a través de sus dibujos de árboles filogenéticos y comparaciones de embriones, que se usaron en textos escolares hasta bien entrado el siglo XX.

Figura 1.34: Ilustración de Haeckel. Imagen disponible bajo licencia Creative Commons CC0.
Además de su impacto en la embriología y la visualización científica, Haeckel tuvo una influencia duradera en la teoría evolutiva y el campo de la biología evolutiva del desarrollo, conocido hoy como Evo-Devo. Aunque su ley biogenética fue ampliamente rechazada, su insistencia en las similitudes embrionarias como evidencia de la evolución dejó un legado duradero. Investigaciones posteriores en embriología comparada han confirmado que los embriones de organismos emparentados muestran notables similitudes durante ciertas etapas de desarrollo, lo que respalda la idea de un ancestro común. De hecho, Haeckel fue uno de los primeros en integrar la embriología dentro del marco de la evolución darwiniana, y su enfoque en los ciclos de vida como un reflejo de la historia evolutiva influyó en generaciones posteriores de biólogos
No obstante, la ley biogenética de Haeckel tuvo una serie de implicaciones problemáticas en su tiempo. Aunque inicialmente resultaba atractiva, su adopción provocó serias repercusiones que afectaron la manera en que se estudiaba la evolución y, especialmente, la paleontología. La teoría de la recapitulación filogenética de Haeckel sugirió que los fósiles no eran esenciales para interpretar las filogenias. En otras palabras, se planteaba que observar las etapas del desarrollo embrionario podía ofrecer una comprensión completa de la historia evolutiva de una especie. Esto generó un distanciamiento de la importancia de los fósiles y del registro paleontológico en la interpretación de las relaciones evolutivas. Aunque la ontogenia aportaba conocimientos valiosos sobre la evolución, se demostró que era insuficiente para entender plenamente la diversidad biológica y los cambios en el mundo natural. La adopción de la recapitulación filogenética afectó negativamente también al desarrollo de enfoques funcionales y ecológicos. Además, las ideas de Haeckel, incluida su hipótesis de la Gastraea, evocaban conceptos de arquetipos similares a los propuestos por Richard Owen, lo que conllevaba profundas implicaciones tanto evolutivas como filosóficas.
Lectura recomendada
Sander, K. (2002). Ernst Haeckel’s ontogenetic recapitulation: irritation and incentive from 1866 to our time. Annals of Anatomy-Anatomischer Anzeiger, 184(6), 523-533.
Watts, E., Levit, G. S., & Hossfeld, U. (2019). Ernst Haeckel’s contribution to Evo-Devo and scientific debate: a re-evaluation of Haeckel’s controversial illustrations in US textbooks in response to creationist accusations. Theory in Biosciences, 138, 9-29.
1.1.20 Finales del siglo XIX: aplicación industrial de la paleontología
A medida que avanzaba el siglo XIX, la paleontología se encontraba en medio de cambios significativos, algunos de los cuales estaban relacionados con su aplicación industrial. Durante el último cuarto del siglo, se comenzó a utilizar el registro fósil en prospecciones de carbón y petróleo (Fig. 1.35), lo que marcó un punto de inflexión en la evolución de esta disciplina.

Figura 1.35: Primer pozo de petróleo en Alberta, en el río Athabasca. Imagen disponible bajo licencia Creative Commons CC0.
Esta aplicación industrial de la paleontología tuvo dos aspectos clave:
Datación y Prospección: Uno de los usos principales de los fósiles en esta etapa era su aplicación en la datación de depósitos geológicos. A través de la identificación de fósiles particulares en las capas de sedimento, los geólogos podían establecer la edad relativa de las formaciones geológicas. Esto era esencial para la industria, especialmente en la búsqueda de recursos como el carbón y el petróleo. Sin embargo, esta aplicación se centraba principalmente en la datación geológica y no en el conocimiento biológico derivado de los fósiles. Se utilizaban para determinar la secuencia de las capas y su antigüedad, proporcionando información útil para la industria pero sin aportar un conocimiento paleobiológico profundo.
Cambio en la Comunicación y Experticia: A medida que la paleontología se incorporaba en la búsqueda de recursos, los “descubrimientos” de fósiles no siempre se comunicaban de manera efectiva. La comunicación de los hallazgos fósiles se volvía más selectiva y, a menudo, se limitaba a expertos en geología y paleontología industrial. Esto tuvo un impacto en la colaboración y la difusión del conocimiento paleontológico en la comunidad científica. Además, esta aplicación industrial también tuvo un impacto en la experticia y las especializaciones dentro de la paleontología. Se produjo una transición en la que los expertos en microfósiles, que eran relevantes para la datación de las capas sedimentarias, ganaron terreno. Los macrofósiles a menudo excedían el tamaño de los tubos de sondeo utilizados en la prospección de recursos y no eran tan útiles para la datación geológica en esta escala. Sin embargo, los expertos en macrofósiles no desaparecieron por completo; en cambio, encontraron su lugar en instituciones de investigación básica, donde su conocimiento continuó contribuyendo a la comprensión de la evolución de organismos más grandes.
1.1.21 Siglo XX: la evolución después de Darwin
El siglo XX fue crucial para el desarrollo de la paleobiología, con Othenio Abel como pionero (Sección 1.1.1.2). Introdujo el término “paleobiología” en Grundzüge der Paläobiologie der Wirbeltiere, proponiendo un enfoque interdisciplinario para entender las adaptaciones y modos de vida de los organismos fósiles.
Casi ocho décadas transcurrieron antes de que la comunidad científica mayoritariamente abrazara la selección natural como el motor principal que da forma a la evolución orgánica, dejando atrás cuatro conceptos alternativos ampliamente aceptados en esa época.
El primer concepto que cuestionaba la selección natural como principal motor de la evolución orgánica era el creacionismo. En su obra seminal, Charles Darwin realizó una afirmación contundente al rechazar esta idea, sosteniendo que las especies no eran el producto de “actos milagrosos de creación”. Sin embargo, a pesar de esta refutación, el creacionismo mantuvo su popularidad en la época de Darwin y continúa vigente en la actualidad, frecuentemente disfrazado bajo el término de “diseño inteligente”.
El segundo concepto que desafió el concepto de la selección natural en la evolución orgánica fue el lamarckismo. Aunque Charles Darwin aceptó un modo de herencia que podría considerarse “lamarckiano” en su trabajo, es importante destacar que su aceptación de la herencia de caracteres adquiridos no fue universal entre sus contemporáneos. Este concepto, a menudo denominado “herencia blanda”, sostenía que los organismos podían adquirir características durante su vida que luego se transmitirían a su descendencia. Darwin incorporó esta idea en su obra, y aunque él mismo creía en la herencia de caracteres adquiridos, no ocurrió lo mismo con Alfred Russel Wallace. Sin embargo, esta perspectiva no se mantuvo universalmente aceptada en la comunidad científica. A pesar de que August Weismann refutó de manera concluyente un efecto directo del entorno en la línea germinal de padres a hijos en los animales en su trabajo de 1892, el lamarckismo siguió siendo popular en algunos círculos hasta alrededor de 1940. Desde la década de 1990, los términos “lamarckismo” y “lamarckiano” han resurgido de manera notable en la literatura biológica. Este resurgimiento ha sido impulsado por el descubrimiento de nuevos mecanismos moleculares (herencia epigenética transgeneracional), como la metilación del ADN, las modificaciones de histonas y la interferencia de ARN, que se han interpretado como evidencia de la existencia y efectividad de la herencia de caracteres adquiridos, lo que ha llevado a un renovado interés en el lamarckismo. Sin embargo, existen dos tipos de argumentos que permiten distinguir claramente entre el concepto genuino lamarckiano de herencia de caracteres adquiridos y la herencia epigenética transgeneracional. El primer argumento se centra en el explicandum de los procesos en cuestión. Los mecanismos moleculares de la herencia epigenética transgeneracional se consideran productos de la evolución a través de la selección natural. Esto implica que el tipo de herencia de caracteres adquiridos que podrían mediar es una característica emergente de la evolución. En contraste, los enfoques tradicionales del lamarckismo ven la herencia de caracteres adquiridos como una propiedad intrínseca de la materia viva. El segundo argumento se refiere al explicans de la herencia de caracteres adquiridos. Según el conocimiento actual, los mecanismos epigenéticos no pueden impulsar la evolución adaptativa de manera independiente. Los fenómenos lamarckianos emergentes solo serían posibles si la variación epigenética individual permitiera que la herencia de caracteres adquiridos fuera un factor de cambio ilimitado, pero no lo es.
El tercer concepto que desafiaba el papel de la selección natural en la evolución orgánica se conoce como ortogénesis. Esta idea fue propuesta en 1893 por el evolucionista alemán William Haacke, quien argumentó que si se permitía cualquier tipo de variación, se esperaría que las reversiones fueran tan frecuentes como las novedades. La rareza de tales reversiones se interpretó como evidencia de que el origen evolutivo de la variación no solo era limitado, sino que también estaba restringido o sesgado por una organización biológica preexistente. La ortogénesis fue adoptada posteriormente por una minoría de evolucionistas, incluidos algunos que la aplicaron a teorías de fuerzas vitales subyacentes o propósitos. Algunos de los defensores de esta idea en su tiempo mantenían una analogía errónea entre la filogenia y la ontogenia, creyendo en una tendencia endógena en la evolución hacia una perfección y complejidad crecientes. Desde esta perspectiva, la evolución era vista como un proceso programado que conduciría finalmente a un resultado predeterminado. Este concepto de ortogénesis sugería que, al igual que en la ontogenia, donde el cigoto se transforma en un embrión y luego en un organismo adulto, existía un mecanismo interno que guiaba hacia productos evolutivos “perfectos”. Sin embargo, estas ideas dejaron de ser tomadas en serio después de la década de 1940, cuando los teóricos darwinianos lograron presentar la ortogénesis como una noción teleológica o mística. A pesar de esto, nuevos desarrollos en genética y biología del desarrollo están dando a conocer nuevos mecanismos que son generalmente consistentes con el concepto original de ortogénesis. Modelos evolutivos como el molecular drive y la mutation-driven evolution, aunque controvertidos, comparten con la ortogénesis la perspectiva de que la selección natural por sí sola no es suficiente ni esencial para entender algunos casos de diferenciación y divergencia evolutivas. Los biólogos evolutivos han reconocido ampliamente que aspectos de la arquitectura genómica, la estructura de los genes y las vías de desarrollo son difíciles de explicar sin involucrar procesos no adaptativos de deriva genética y mutación. Complementario a los estudios genéticos y de desarrollo, existe una tendencia generalizada en la morfología que involucra la reducción, supresión y fusión en estructuras complejas multipartitas, como la flor de angiosperma y el cráneo de los vertebrados.
El cuarto concepto que desafió la primacía de la selección natural en la evolución orgánica fue el transmutacionismo, específicamente la variante saltacionista de esta idea. Esta teoría, influenciada por el pensamiento tipológico, sostenía que un organismo podía transformarse en una forma de vida sustancialmente diferente, a menudo en una o dos generaciones, debido a lo que se llamaba “macromutaciones”. Los defensores de esta perspectiva, como el genetista Goldschmidt, reconocían que la mayoría de las macromutaciones producirían “monstruos” inviables, pero argumentaban que ocasionalmente surgirían “monstruos prometedores” que estarían bien adaptados a un nuevo entorno. Esta idea sugería que un nuevo tipo de organismo, o incluso un linaje completo, podría evolucionar sin depender de la selección natural. Sin embargo, esta teoría fue refutada a medida que se entendía mejor la genética y la biología. Se hizo evidente que los organismos no son tipos estáticos, como moléculas de glucosa, sino que las poblaciones están compuestas por numerosas variantes genómicas. Por lo tanto, la aparición de un solo “monstruo prometedor” no sería suficiente para impulsar la evolución, ya que necesitaría reproducirse con otro “monstruo prometedor” del sexo opuesto. Aunque raramente se han observado macromutaciones que tengan efectos fenotípicos significativos, como sugieren los transmutacionistas, algunas investigaciones indican que ciertas mutaciones con grandes efectos fenotípicos pueden haber desempeñado un papel en la evolución de los invertebrados. Sin embargo, la teoría original de los “monstruos prometedores”, tal como la concebían Goldschmidt y otros, no cuenta con respaldo experimental sólido. El problema central no es si los “monstruos prometedores” jugaron algún papel en la evolución orgánica, sino si representan el mecanismo predominante detrás de los cambios evolutivos radicales.
Lectura recomendada
Dietrich, M. R. (2003). Richard Goldschmidt: hopeful monsters and other’heresies’. Nature Reviews Genetics, 4(1), 68-74.
Grehan, J. R. (2018). Orthogenesis. The International Encyclopedia of Biological Anthropology, 1-2.
Loison, L. (2018). Lamarckism and epigenetic inheritance: a clarification. Biology & Philosophy, 33, 1-17.
1.1.22 Siglo XX: la evolución biológica a lo largo de las eras: tres etapas clave en el desarrollo de la Teoría Moderna.
El desarrollo de la teoría moderna de la evolución se puede dividir en tres etapas distintas que marcaron avances significativos en nuestra comprensión de cómo se lleva a cabo la evolución biológica (Fig. 1.36).
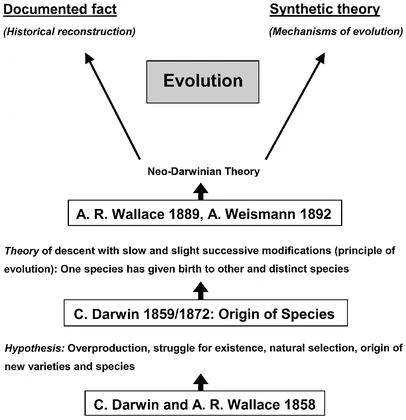
Figura 1.36: Esquema que ilustra el desarrollo histórico del concepto de evolución: desde la hipótesis de Darwin y Wallace, pasando por Darwin, Wallace y Weismann, hasta la teoría sintética. Imagen tomada de Kutschera & Niklas (2004).
1.1.22.1 Darwinismo
Esta etapa se inicia con la publicación de “El Origen de las Especies” por Charles Darwin en 1859, un hito crucial en la historia de la biología. En su obra, Darwin formuló el principio de la selección natural, que también fue propuesto de manera independiente por Alfred Russel Wallace. Esta idea fundamental sostiene que las especies evolucionan a lo largo del tiempo a través de la adaptación de individuos mejor adaptados a su entorno y la transmisión de sus características a las generaciones futuras. Un aspecto interesante de esta etapa es que Darwin aceptó, en su momento, el principio de Lamarck sobre la herencia de las características adquiridas como una fuente de variabilidad biológica. Esto llevó a un período de pensamiento evolutivo denominado “Lamarck/Darwin/Wallace”. En 1872, se publica la sexta edición de “El Origen de las Especies”, donde Darwin continuó refinando y expandiendo sus ideas originales.
1.1.22.2 Neo-Darwinismo
La etapa del Neo-Darwinismo se caracterizó por avances clave en la comprensión de la variabilidad biológica. August Weismann, en 1892, proporcionó evidencia experimental en contra de la herencia “Lamarckiana”. Postuló que la reproducción sexual, que involucra la recombinación genética, crea en cada generación una población nueva y variable de individuos. Alfred Russel Wallace, quien popularizó el término “darwinismo”, incorporó plenamente las conclusiones novedosas de Weismann y se convirtió en uno de los primeros defensores del neodarwinismo. El término “neodarwinismo” se refiere a una teoría expandida de Darwin que excluye la herencia lamarckiana como un mecanismo viable para la evolución.
1.1.22.3 Síntesis
La etapa de la Síntesis, que tuvo lugar entre 1937 y 1950, marcó un período de avances significativos al incorporar hallazgos de diversas disciplinas científicas, como la genética, la sistemática y la paleontología. A menudo se denomina “teoría sintética” o “síntesis neodarwiniana”, esta etapa unificó y consolidó las ideas evolutivas en una estructura coherente. La teoría sintética integra la selección natural de Darwin con los conceptos genéticos de Mendel y los hallazgos fósiles de la paleontología, proporcionando una comprensión más completa de cómo evolucionan las especies.
Lectura recomendada
Kutschera, U., & Niklas, K. J. (2004). The modern theory of biological evolution: an expanded synthesis. Naturwissenschaften, 91(6), 255-276.
1.1.23 La síntesis evolutiva: los principios fundamentales de la teoría moderna de la evolución
El siglo XX marcó un hito en el desarrollo de la teoría de la evolución con la llegada de la “Síntesis Evolutiva”. Este proceso intelectual, que tuvo lugar entre 1930 y 1950, involucró a un grupo diverso de biólogos de varios países. Su objetivo era unificar las ideas sobre la evolución y consolidarlas en una teoría coherente. Como resultado, se creó un conjunto de declaraciones de consenso que constituyen el núcleo de la teoría sintética o moderna de la evolución biológica.
Los arquitectos de la teoría moderna, como el naturalista/genetista ruso/estadounidense Theodosius Dobzhansky (1900–1975), el naturalista/sistematista alemán/estadounidense Ernst Mayr (1904–2005), el zoólogo británico Julian Huxley (1887–1975), el paleontólogo estadounidense George G. Simpson (1902–1984), el zoólogo alemán Bernhard Rensch (1900–1990) y el botánico estadounidense G. Ledyard Stebbins (1906– 2000), contribuyeron significativamente a este proyecto intelectual. A través de sus esfuerzos y obras, se establecieron seis principios fundamentales que han dado forma a nuestra comprensión actual de la evolución (Fig. 1.37):
Poblaciones como Unidades de Evolución: La teoría sintética postula que las unidades de evolución son poblaciones de organismos, en contraposición a la idea de tipos fijos. En otras palabras, la evolución opera a nivel de poblaciones y no en individuos aislados.
Variabilidad Genética y Fenotípica: La variabilidad genética y fenotípica en las poblaciones de plantas y animales se debe a la recombinación genética. Esta recombinación resulta de la reproducción sexual y mutaciones aleatorias a lo largo de la secuencia de padres e hijos. La diversidad genética es la base de la evolución.
Selección Natural: La selección natural se reconoció como la fuerza más importante que da forma al curso de la evolución fenotípica. La idea central es que los individuos con características beneficiosas tienen una ventaja en la supervivencia y la reproducción, lo que conduce a la propagación de esas características en la población. Además, en poblaciones pequeñas, la deriva genética puede ser significativa.
Especiación: La especiación se define como un paso crucial en el proceso evolutivo en el que las formas se vuelven incapaces de cruzarse entre sí. Se han propuesto varios mecanismos de aislamientos (Sección @ref(Especies-y-especiación)).
Transiciones Evolutivas Graduales: En la teoría sintética, se reconoce que las transiciones evolutivas en las poblaciones, tanto en términos de cambios alélicos como fenotípicos, tienden a ser graduales. Si bien la inmigración de individuos de poblaciones vecinas no puede ser ignorada, se considera de menor importancia en comparación con la selección natural.
Macroevolución: La macroevolución, que se refiere a cambios evolutivos por encima del nivel de especie, se entiende como un proceso gradual paso a paso. Este concepto no es más que una extrapolación de la microevolución, lo que significa que los mismos principios evolutivos que se aplican a nivel de población también se aplican a la evolución a gran escala.
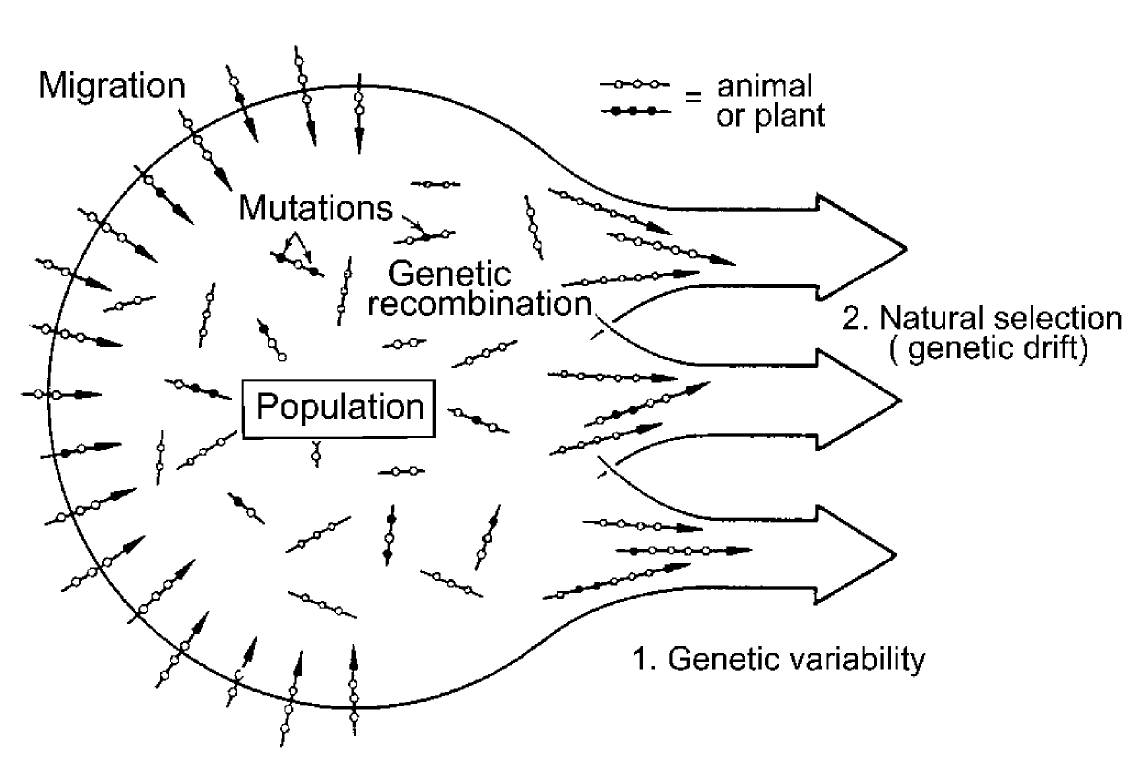
Figura 1.37: Esquema para ilustrar la interacción de los procesos básicos que provocan la evolución fenotípica en una población variable de organismos (animales o plantas). Imagen tomada de Kutschera & Niklas (2004).
La teoría sintética de la evolución consolida estos principios en una estructura coherente, brindando una comprensión profunda de cómo la diversidad de la vida en la Tierra ha evolucionado a lo largo del tiempo. Como resultado, esta teoría se ha convertido en una base fundamental para la biología evolutiva y sigue siendo un pilar crucial en la investigación científica. La síntesis evolutiva nos ha proporcionado una visión más profunda de la asombrosa diversidad de la vida en nuestro planeta y ha revolucionado nuestra comprensión de cómo todas las formas de vida están interconectadas a través de procesos evolutivos comunes.

1.1.23.0.1 Fuerzas evolutivas: La selección natural y la deriva genética
La selección natural es un mecanismo evolutivo clave que opera sobre la variación fenotípica dentro de una población, favoreciendo la proliferación de individuos cuyas características confieren una ventaja adaptativa en un entorno particular. A través de este proceso diferencial de supervivencia y reproducción, los alelos asociados a estos rasgos beneficiosos incrementan su frecuencia a lo largo de las generaciones, lo que resulta en una mayor adaptación de la población al entorno. Este mecanismo, basado en la interacción entre los fenotipos y las presiones ambientales, constituye la principal fuerza impulsora de la adaptación biológica.
La deriva genética es una fuerza evolutiva que cambia aleatoriamente las frecuencias alélicas en una población a lo largo del tiempo. A diferencia de la selección natural, no depende de la adaptación al entorno, sino de procesos aleatorios, como el muestreo en la reproducción. Este fenómeno es más pronunciado en poblaciones pequeñas, donde puede provocar la pérdida de alelos raros y la fijación de alelos comunes, reduciendo la diversidad genética. El efecto de cuello de botella (una reducción drástica en la población) o el efecto fundador (el establecimiento de una nueva población a partir de un pequeño grupo) pueden amplificar su impacto. En poblaciones grandes, su influencia es menor y las frecuencias alélicas tienden a estabilizarse.
1.1.24 Siglo XX: avances en las subdisciplinas de la paleontología
El siglo XX trajo consigo un período de gran expansión y desarrollo en las subdisciplinas de la paleontología. Mientras la paleontología clásica se centraba principalmente en la identificación y descripción de fósiles, el nuevo siglo vio surgir un interés renovado en campos especializados que arrojaron luz sobre la historia de la vida en la Tierra de maneras más profundas y completas. Aquí, destacaremos tres de estas subdisciplinas que ganaron prominencia en el siglo XX: la tafonomía, la paleoecología y la paleobiogeografía.
1.1.24.1 Tafonomía: descifrando el proceso de fosilización
En la década de 1940, Ivan Efremov (1908-1972), un paleontólogo ruso, desempeñó un papel fundamental en el desarrollo de la tafonomía, una rama de la paleontología que se centra en comprender los procesos que afectan a los restos orgánicos después de la muerte de un organismo y cómo se convierten en fósiles. La tafonomía ha demostrado ser esencial para interpretar y comprender el registro fósil, ya que arroja luz sobre cómo los restos orgánicos se preservan y alteran con el tiempo. El estudio de la tafonomía ha revelado información valiosa sobre la fosilización, la formación de yacimientos fósiles y cómo los restos se degradan y se convierten en fósiles.
1.1.24.2 Paleoecología: explorando ecosistemas del pasado
La paleoecología, una disciplina que se enfoca en la reconstrucción de los ecosistemas del pasado y las interacciones entre las especies, experimentó un resurgimiento en las décadas de 1930 y 1940. Paleontólogos rusos y alemanes jugaron un papel destacado en revitalizar el interés por esta disciplina. Fue en este período que se acuñó el término “paleoecología”, y se realizaron síntesis significativas en las décadas de 1960 y 1970. La paleoecología no solo nos permite comprender cómo eran los ecosistemas del pasado, sino que también proporciona información crucial sobre la evolución y las adaptaciones de las especies en respuesta a cambios ambientales a lo largo del tiempo.
1.1.24.3 Paleobiogeografía: en el contexto de la tectónica de placas
La paleobiogeografía, que se enfoca en el estudio de la distribución geográfica de las especies extintas a lo largo de la historia de la Tierra, experimentó un cambio significativo en la década de 1970. Esto se debió en gran parte al surgimiento de la teoría de la tectónica de placas, que revolucionó nuestra comprensión de la dinámica de la Tierra. La teoría de la tectónica de placas explicó cómo los continentes se han movido a lo largo de millones de años y cómo estos movimientos han influido en la distribución de las especies. La paleobiogeografía se benefició enormemente de esta teoría, ya que ahora podía comprender cómo los patrones de distribución de los fósiles estaban relacionados con la deriva continental y otros cambios geológicos.
1.1.25 Síntesis evolutiva en el siglo XX: entre el estancamiento y la renovación
En la década de 1950, la síntesis evolutiva comenzó a mostrar signos de estancamiento. En lugar de avanzar en todas las direcciones posibles, la atención se centró principalmente en la selección natural y la adaptación como las fuerzas motrices principales en la evolución. La creencia predominante sostenía que la selección natural produciría tendencias evolutivas que, en última instancia, conducirían a una mejora adaptativa en las especies. Esta perspectiva llevó a una especie de tautología adaptacionista, en la que cualquier tendencia evolutiva se interpretaba en términos de mejora por selección, lo que limitaba la exploración de otras fuerzas y mecanismos que podrían impulsar la evolución.
Una de las razones detrás de este anquilosamiento fue la falta de comprensión sobre el peso adaptativo de muchos cambios morfológicos. En otras palabras, no se sabía con certeza cuánto contribuían estos cambios a la supervivencia y reproducción de las especies. Esta incertidumbre perpetuaba la tautología adaptacionista, ya que la falta de datos concretos sobre la adaptación de los rasgos morfológicos dificultaba la consideración de otras influencias en la evolución.
1.1.25.1 El método de los paradigmas

Figura 1.38: Retrato de Martin Rudwick. © The University of Chicago Press.
A medida que el siglo XX avanzaba, se produjo un cambio significativo en el campo de la paleobiología, impulsado por la visión innovadora del paleontólogo británico Martin Rudwick (1932-Actualidad) (Fig. 1.38). Rudwick reconoció esta limitación fundamental en la síntesis evolutiva, que en gran medida se centraba en la selección natural y la adaptación, y que a menudo carecía de un análisis detallado de las estructuras morfológicas y su función en organismos fósiles. Para abordar esta deficiencia, Rudwick propuso una revolución morfofuncional, introduciendo el método de los “paradigmas” como una herramienta fundamental.
La idea detrás de esta revolución morfofuncional era investigar la mecánica del funcionamiento de las estructuras biológicas en organismos fósiles para inferir su función y optimalidad. Rudwick adoptó un enfoque que se basó en el análisis funcional de estas estructuras, buscando pistas en su morfología que permitieran deducir su función. El concepto de “paradigmas” se convirtió en el núcleo de su enfoque, que implicaba que una estructura morfológica debería ser capaz de desempeñar su función con eficiencia. La analogía con una tetera ayudó a ilustrar este principio: al igual que alguien no familiarizado con el consumo de té podría deducir la función del pico de una tetera examinando su estructura y diseño, Rudwick sostenía que este enfoque podía aplicarse a organismos fósiles.
Rudwick argumentó que si se encontrara que la morfología de una estructura no era adecuada para desempeñar una función eficiente, entonces podría descartarse esa función en particular como improbable. Tomando la analogía de la tetera, si se demostrara que el pico no era adecuado para verter té de manera eficiente, se podrían explorar interpretaciones alternativas de su función, o incluso concluir que no tenía función en absoluto. Este enfoque no presupone que todas las características de un organismo deban tener un valor funcional o adaptativo en términos físicos; más bien, busca probar y evaluar las interpretaciones funcionales en función de principios físicos o mecánicos relevantes. Por tanto, el término “paradigma” en el contexto de Rudwick deriva de la idea de características funcionales “idealmente eficientes”, lo que coincide con su etimología en el sentido de ser un “modelo” o un “ejemplo claro y típico” de cómo debería funcionar una estructura de manera óptima.
La revolución morfofuncional de Rudwick trajo consigo una nueva perspectiva para la paleobiología al enfocarse en las estructuras anatómicas de los organismos fósiles y su funcionalidad. Esta aproximación permitió un análisis más profundo de la biología de los seres extintos, y aunque no fue inmune a la crítica y el debate, sirvió como un paso importante hacia la ruptura de la tautología adaptacionista. La visión de Rudwick abrió nuevas posibilidades para explorar las relaciones entre la morfología y la función en el contexto de la evolución biológica, y dejó una marca duradera en la paleobiología y la interpretación del registro fósil. Su enfoque en la inferencia funcional y el análisis morfológico sigue siendo relevante y sigue siendo un componente valioso en la paleobiología moderna.
Lectura recomendada
Rudwick, M. J. (2018). Functional Morphology in Paleobiology: Origins of the Method of ‘Paradigms’. Journal of the History of Biology, 51(1), 135-178.
1.1.25.2 Biomorfodinámica y las limitaciones evolutivas: el legado de Adolf Seilacher

Figura 1.39: Retrato de Adolf Seilacher. Imagen disponible bajo licencia Creative Commons CC0.
Adolf Seilacher (1925-2014) (Fig. 1.39), un influyente paleontólogo alemán, hizo varias contribuciones significativas a la paleontología a lo largo de su carrera. Entre sus propuestas destacadas se encuentra la noción de morfología construccional, también conocida como biomorfodinámica, que ofrece una perspectiva única sobre las limitaciones que influyen en el cambio evolutivo. Estas limitaciones se pueden dividir en tres categorías fundamentales: restricciones funcionales/adaptativas, restricciones históricas/filogenéticas y restricciones constructivas/morfogenéticas (Fig. 1.40). La morfología construccional de Seilacher aporta una perspectiva valiosa al abordar las limitaciones del enfoque adaptacionista en la síntesis evolutiva permitiendo un análisis más amplio de los factores que modelan la diversidad biológica.
Figura 1.40: Visualización original de la morfología construccional de Seilacher. Imagen disponible bajo licencia Creative Commons CC0.
Restricciones Funcionales/Adaptativas: o Estas restricciones se relacionan con las demandas funcionales y adaptativas que los organismos enfrentan en sus respectivos entornos. Los seres vivos deben ajustar su morfología y características físicas para cumplir con ciertas funciones, como la alimentación, la locomoción y la reproducción. o La selección natural juega un papel crucial en la imposición de restricciones funcionales. Los individuos que no pueden realizar estas funciones de manera eficiente o adaptativa tienden a tener menos éxito reproductivo, lo que reduce la probabilidad de transmitir sus rasgos a las generaciones futuras. o Las restricciones funcionales son evidentes en la forma en que los organismos evolucionan para satisfacer las demandas de su nicho ecológico. Por ejemplo, las aves desarrollaron alas adaptadas para el vuelo, lo que limita las modificaciones extremas en la forma de sus extremidades anteriores.
Restricciones Históricas/Filogenéticas: o Estas restricciones se derivan de la historia evolutiva de un linaje particular. Las características heredadas de los ancestros pueden imponer limitaciones a la dirección en la que evolucionará un grupo de organismos. o La historia filogenética de un organismo a menudo deja una huella en su morfología. Algunas características que no se relacionan directamente con la adaptación a su entorno pueden ser conservadas debido a su herencia de un ancestro común. o Las restricciones históricas pueden influir en la evolución en ciertas direcciones predecibles. Por ejemplo, los mamíferos tienen una estructura ósea particular en sus extremidades debido a la herencia de un ancestro tetrapodo.
Restricciones Constructivas/Morfogenéticas: o Estas restricciones se centran en las limitaciones impuestas por los procesos de desarrollo y crecimiento de los organismos. Los patrones de desarrollo, como la formación de estructuras óseas y musculares, pueden restringir las formas que pueden adquirir. o La genética y los mecanismos de desarrollo establecen límites en la morfología de un organismo. Las mutaciones y los cambios genéticos pueden conducir a variaciones morfológicas, pero dentro de ciertos límites. o Las restricciones constructivas se relacionan con la viabilidad de una estructura o forma en un organismo. Si una modificación morfológica excede las capacidades de desarrollo de un organismo o no es compatible con su supervivencia, es poco probable que ocurra.
Lectura recomendada
Tamborini, M. (2020). Challenging the Adaptationist Paradigm: Morphogenesis, Constraints, and Constructions. Journal of the History of Biology, 53(2), 269-294.
1.1.26 Equilibrio interrumpido o puntuado en la teoría de la evolución

Figura 1.41: Retratos de Stephen Jay Gould (izquierda) y Niles Eldredge (derecha). © timetoast.
El siglo XX fue testigo de un cambio significativo en la teoría de la evolución, impulsado por las observaciones en el registro fósil que desafiaban las expectativas de la síntesis evolutiva moderna. El equilibrio interrumpido o puntuado, una teoría propuesta por los paleontólogos Niles Eldredge y Stephen Jay Gould en 1972 (Fig. 1.41), plantea que la evolución no siempre ocurre de manera gradual y constante, como se había asumido previamente. Este cambio de paradigma en la paleontología tuvo implicaciones importantes en nuestra comprensión de la evolución biológica y marcó una distinción entre la macroevolución y la microevolución.
La teoría del equilibrio interrumpido postula que, una vez que se forma una nueva especie, esta tiende a mantener su morfología estable durante largos períodos de tiempo. En otras palabras, las especies son morfológicamente estables durante la mayor parte de su historia evolutiva. Este concepto desafió la noción de una evolución constante y gradual propuesta por la síntesis evolutiva moderna, que asumía que los cambios morfológicos ocurrían de manera continua y acumulativa en el tiempo.
Eldredge y Gould argumentaron que la estabilidad morfológica se ve interrumpida solo en eventos de especiación. Durante estos episodios de especiación, nuevas especies se originan y muestran cambios significativos en su morfología. Sin embargo, una vez que se establece una nueva especie, su morfología tiende a estabilizarse nuevamente. Esto implica que la evolución morfológica es “puntuada” por episodios de cambio en lugar de ser un proceso constante (Fig. 1.42).
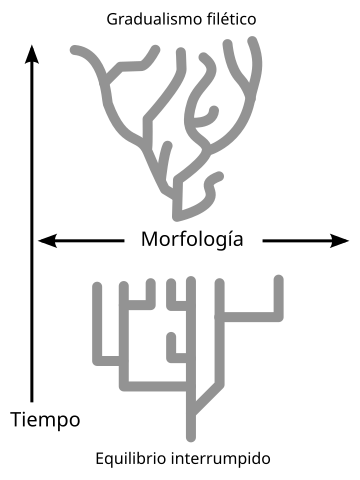
Figura 1.42: Diagrama que ejemplifica el gradualismo filético (arriba) y el equilibrio interrumpido (abajo). Imagen disponible bajo licencia Creative Commons CC0.
El concepto de equilibrio interrumpido tiene varias implicaciones significativas:
Desacople entre Macro- y Microevolución: El equilibrio puntuado propone un desacople entre la macroevolución y la microevolución, subrayando la complejidad de la evolución a diferentes niveles. La microevolución se centra en los cambios en las frecuencias alélicas dentro de las poblaciones a lo largo del tiempo, mientras que la macroevolución aborda las transformaciones que ocurren a nivel de especies y grupos taxonómicos superiores. Bajo esta perspectiva, la macroevolución ya no se considera simplemente una extrapolación de los procesos microevolutivos, sino que se reconoce como un fenómeno más complejo que puede incluir episodios de estabilidad seguidos de cambios rápidos.
Limitaciones de la Síntesis Evolutiva: El equilibrio puntuado plantea un desafío a la síntesis evolutiva moderna, que se centraba en gran medida en la selección natural y los cambios graduales en las poblaciones. La idea de que las especies son morfológicamente estables durante la mayor parte de su historia destaca que la evolución es un proceso más complejo de lo que se había asumido previamente.
1.1.27 La diversidad fanerozoica y las extinciones en masa

Figura 1.43: Jack Sepkoski (izquierda) con David Raup (derecha). © extinctblog
Durante el siglo XX, los científicos comenzaron a investigar y documentar la historia de la diversidad biológica a través de la paleontología y el análisis de fósiles. A medida que avanzaban en su comprensión de la diversidad, surgió una pregunta fundamental: ¿qué causó los patrones de diversidad, extinción y originación a lo largo del fanerozoico? En la década de 1980, se produjo un enfoque importante en el estudio de la diversidad y las extinciones en masa y sus mecanismos. Los científicos se centraron exclusivamente en los factores ambientales que pudieron haber contribuido a estos patrones.
Uno de los investigadores clave en este campo fue David M. Raup (1933-2015) (Fig. 1.43), cuyos trabajos se centraron en modelar el juego de la originación y la extinción en la historia de la vida en la Tierra. Raup examinó cómo los cambios en el ambiente terrestre y marino, como la glaciación, el vulcanismo masivo y el impacto de asteroides, podrían haber desencadenado extinciones a gran escala. Sus modelos ayudaron a arrojar luz sobre cómo los cambios ambientales afectaron la biodiversidad y cómo las especies respondieron a estos eventos catastróficos.
Jack Sepkoski (1948-1999) (Fig. 1.43), otro paleontólogo influyente, contribuyó significativamente a la comprensión de las extinciones en masa y los patrones de diversificación biológica a lo largo del fanerozoico. Su trabajo proporcionó una visión más clara de cómo las extinciones se correlacionaron con eventos geológicos y climáticos, lo que ayudó a revelar la complejidad de los mecanismos detrás de estas extinciones.
Lectura recomendada
Sepkoski, J. J. (1997). Biodiversity: past, present, and future. Journal of Paleontology, 71(4), 533-539.
1.1.28 Posturas internlistas: la criticalidad autoorganizada

Figura 1.44: Ricard Solé. Imagen disponible bajo licencia Creative Commons CC0.
La criticalidad autoorganizada, un concepto que se ha aplicado en el campo de sistemas complejos, ha revolucionado nuestra comprensión de estos sistemas en el siglo XX. El científico Ricard Solé (1962-Actualidad) (Fig. 1.44) ha desempeñado un papel fundamental en la aplicación de este concepto en diversos campos de la biología evolutiva. La criticalidad autoorganizada sostiene que los sistemas con un gran número de componentes pueden evolucionar hacia un estado crítico, en el cual una pequeña perturbación puede provocar una reacción en cadena de proporciones gigantescas.
La criticalidad autoorganizada nos lleva al límite entre el orden y el caos. Los sistemas en estado crítico son inherentemente inestables pero autorregulados, y cualquier pequeña perturbación puede desencadenar un efecto dominó de gran alcance. La criticalidad autoorganizada se puede ilustrar con el ejemplo de un montículo de arena. En este sistema, se añaden lentamente granos de arena a una pila. A medida que se agregan más granos, la pendiente de la pila se vuelve cada vez más inestable. Una vez que se alcanza una inclinación crítica, la adición de un solo grano de arena puede provocar una avalancha que involucra varios granos de arena. En la mayoría de los casos, estas avalanchas son pequeñas y afectan solo a unos pocos granos. Sin embargo, ocasionalmente, se producen avalanchas mucho más grandes que pueden movilizar una gran cantidad de arena. Lo interesante de este sistema es que la frecuencia de las avalanchas de un tamaño determinado, N(s), donde s representa el número de granos involucrados, sigue una ley de potencias. Esto significa que existe una relación matemática que describe cómo la probabilidad de que ocurran avalanchas de diferentes tamaños está distribuida de manera específica, con muchas pequeñas avalanchas y pocas grandes avalanchas (Fig. 1.45). Este fenómeno es característico de sistemas que exhiben criticalidad autoorganizada, donde el comportamiento emergente del sistema no es evidente en los componentes individuales, sino que surge de la interacción colectiva de muchos elementos.
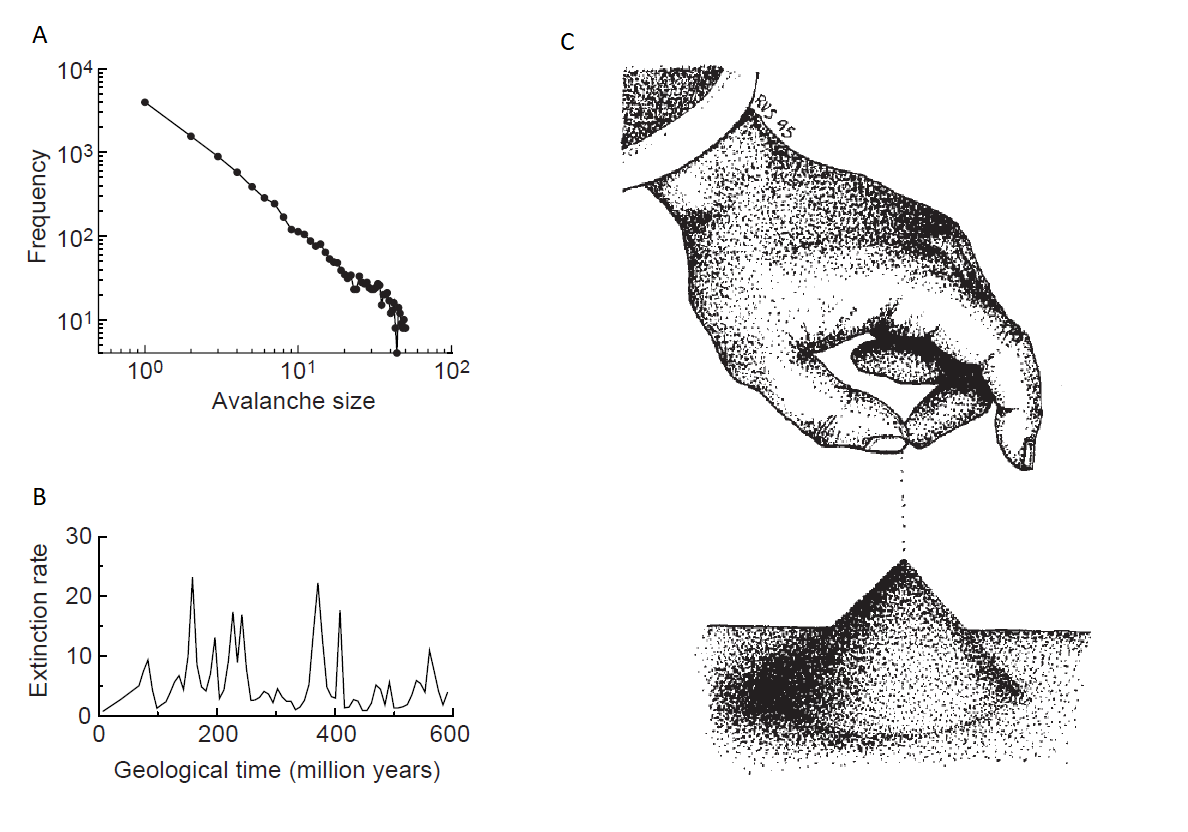
Figura 1.45: Escalado y autosimilitud. (A) Ley de potencias: en un gráfico log-log, una distribución de ley de potencias presenta una línea recta. Se observan muchos eventos pequeños, pero también hay una probabilidad de que ocurran eventos grandes. (B) Fluctuaciones temporales en el registro fósil: se muestra la tasa total de extinción (3 x 100) de familias de animales marinos durante el Fanerozoico, revelando un amplio espectro de eventos de extinción. (C) Ejemplo clasico de criticalidad autoorganizada, el montículo de arena. Imagen tomada de Solé et al. (1999)
Sistemas muy diversos parecen ajustarse a esta relación matematica, encontrándose en una amplia gama de fenómenos naturales y sociales, desde terremotos hasta mercados financieros y ecosistemas. En el campo de la sismología, la criticalidad autoorganizada ha ayudado a explicar la dinámica de los terremotos. Los sistemas geológicos tienden a evolucionar hacia un estado crítico, donde incluso un evento menor puede desencadenar un terremoto significativo. Esta perspectiva ha mejorado nuestra comprensión de la actividad sísmica y ha arrojado luz sobre la geología. La ecología también se ha beneficiado de la teoría de la criticalidad autoorganizada. Los ecosistemas son sistemas complejos en los que las especies están interconectadas. A medida que las conexiones entre especies aumentan, el sistema evoluciona hacia un estado crítico. La rotura de algunas de estas conexiones, como ocurre en eventos de extinción en masa, puede provocar cambios drásticos en la estructura del ecosistema. Un ejemplo notorio es la extinción masiva del Pérmico, donde cambios climáticos y actividad volcánica en Siberia se han atribuido a la extinción. Sin embargo, la teoría de la criticalidad autoorganizada sugiere que la rotura de conexiones entre especies, más que las causas ambientales, desempeñó un papel importante en la extinción de una gran parte de la biota. Esto incorpora una perspectiva internalista para entender las dinámicas de diversidad y extinción en el fanerozoico, en contraposición a las visiones de Raup y Sepkosky centradas exclusivamente en mecanismos ambientales.
Lectura recomendada
Bak, P., & Chen, K. (1991). Criticalidad auto-organizada. Investigación y Ciencia, (174), 18-25.
Solé, R. V., Manrubia, S. C., Benton, M., Kauffman, S., & Bak, P. (1999). Criticality and scaling in evolutionary ecology. Trends in ecology & evolution, 14(4), 156-160.
1.1.29 Siglo XXI ¿Es necesaria una nueva síntesis?
El siglo XXI ha visto surgir debates cruciales sobre la necesidad de una nueva síntesis evolutiva. La síntesis moderna, que se desarrolló entre 1918 y 1942, proporcionó un marco sólido para comprender la evolución. Sin embargo, en las últimas décadas, se ha argumentado que esta síntesis requiere una revisión y expansión significativa para abordar cuestiones más complejas y contemporáneas en biología evolutiva.
La propuesta de una “síntesis evolutiva extendida” se basa en la idea de que los conceptos teóricos previos son insuficientes para abordar ciertos aspectos de la evolución. Esta idea ha evolucionado con el tiempo y ha sido respaldada por varios científicos destacados. A principios de la década de 1950, C. H. Waddington sugirió la necesidad de una síntesis evolutiva más amplia. En la década de 1980, Stephen Jay Gould y Niles Eldredge introdujeron el concepto de “equilibrio puntuado”, que desafió las nociones previas de cambios evolutivos graduales y lentos.
La síntesis evolutiva extendida se basa en una revisión crítica de la síntesis moderna y agrega una serie de conceptos teóricos complementarios para abordar las complejidades emergentes en la biología evolutiva. Algunos de estos conceptos incluyen:
Selección multinivel: Reconoce que la selección natural puede operar en diferentes niveles, desde el nivel genético hasta el nivel del grupo.
Plasticidad fenotípica: Considera la capacidad de los organismos para cambiar su fenotipo en respuesta a cambios en el entorno.
Herencia epigenética transgeneracional: Explora cómo las modificaciones epigenéticas pueden transmitirse a través de generaciones y afectar la evolución.
Construcción de nichos: Examina cómo los organismos pueden modificar su entorno y, a su vez, ser moldeados por él.
Evolucionabilidad: Se refiere a la capacidad inherente de los organismos para evolucionar y adaptarse.
Conceptos de la biología evolutiva del desarrollo (evo-devo): Explora cómo los procesos de desarrollo de un organismo influyen en su evolución.
Sin embargo, no hay un acuerdo general sobre la necesidad de una nueva síntesis evolutiva. Los opositores argumentan que la síntesis moderna es suficiente para explicar los fenómenos recientes observados en la biología evolutiva. Por otro lado, los defensores de la síntesis evolutiva extendida argumentan que las concepciones centrales de la síntesis moderna son demasiado restrictivas y no pueden abordar completamente las observaciones contemporáneas.
En última instancia, la discusión en curso sobre la necesidad de una nueva síntesis evolutiva refleja la dinámica de la ciencia, donde las teorías evolucionan para adaptarse a un creciente cuerpo de evidencia y una comprensión en constante evolución de la biología evolutiva. La resolución de este debate tendrá implicaciones significativas para el futuro de la biología y nuestra comprensión de la evolución.
Lectura recomendada
Laland, K., Uller, T., Feldman, M., Sterelny, K., Müller, G. B., Moczek, A., … & Strassmann, J. E. (2014). Does evolutionary theory need a rethink?. Nature, 514(7521), 161-164.
1.2 La naturaleza del registro fósil
1.2.1 Distribución geográfica y ambiental de las rocas fosilíferas
La distribución geográfica y ambiental de las rocas fosilíferas es esencial para comprender el registro fósil a lo largo del tiempo geológico. Esta distribución varía significativamente debido a varios factores clave. Un patrón importante es que la exposición de rocas con fósiles aumenta de forma exponencial hacia edades geológicas más recientes (Fig. 1.46). Esto se debe a procesos geológicos como la erosión y la tectónica de placas, lo que hace que sea más probable encontrar afloramientos de rocas con fósiles de épocas más recientes en comparación con las más antiguas. Por otro lado, los registros fósiles marinos son más abundantes que los terrestres, principalmente debido a tasas de sedimentación más altas en ambientes marinos en comparación con ambientes terrestres, donde la erosión es común. Además, los registros de ambientes marinos someros superan en número a los de ambientes marinos profundos, ya que los sedimentos someros tienen tasas de acumulación más altas y, por lo tanto, tienen más probabilidades de conservar fósiles a lo largo del tiempo geológico. Sin embargo, en algunas áreas, como zonas con alta erosión, como regiones montañosas de elevada altitud, los registros fósiles son raros o están ausentes debido a la destrucción y erosión de las rocas sedimentarias que contienen fósiles.
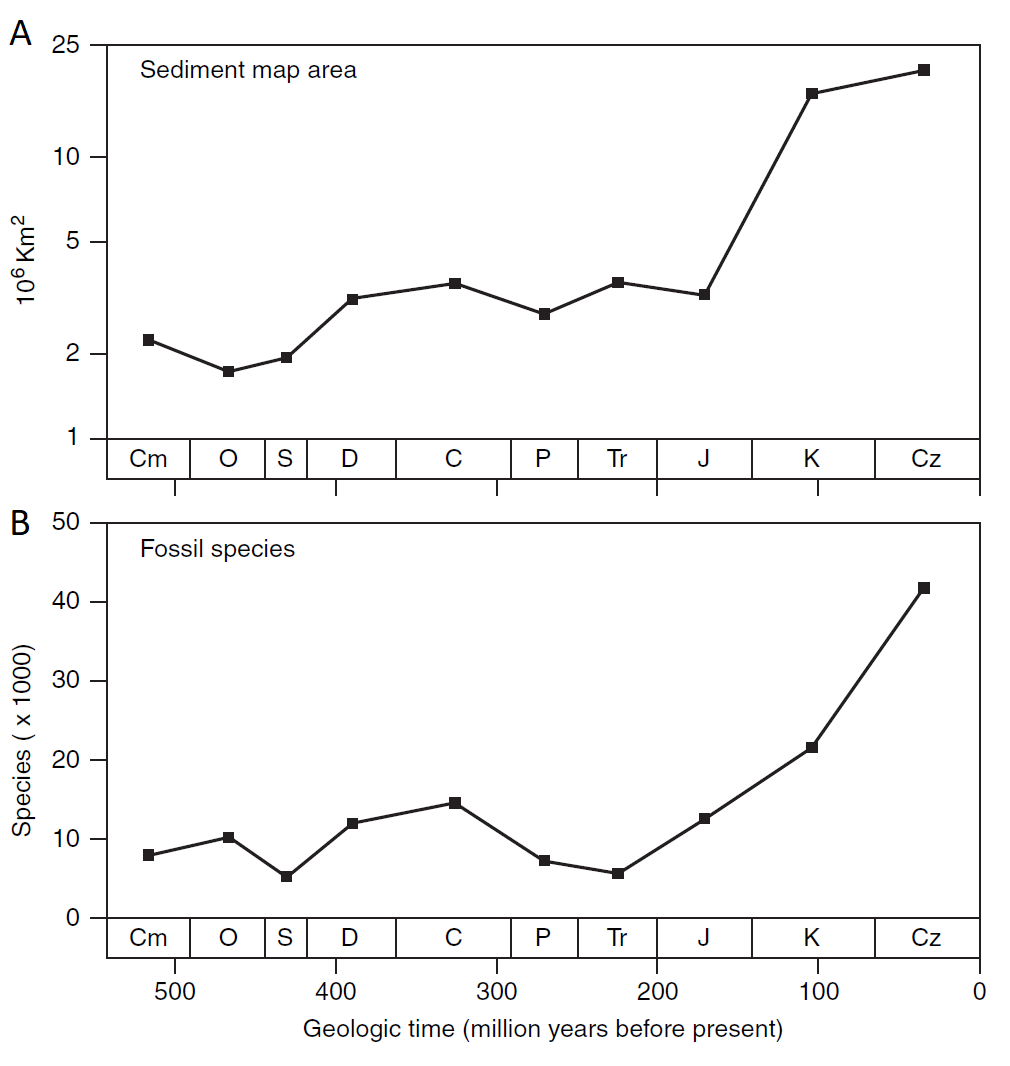
Figura 1.46: Rocas sedimentarias y diversidad fósil a lo largo del tiempo geológico. (A) Área de afloramiento de rocas sedimentarias según mapas geológicos para diferentes periodos del Fanerozoico. Los datos se presentan en una escala logarítmica para resaltar la variación proporcional entre los números. Una diferencia de unidad en esta escala representa una relación constante; por ejemplo, la diferencia entre 1 y 2 es la misma que entre 5 y 10. (B) Estimación del número de especies de invertebrados descubiertas y nombradas entre 1900 y 1975. Los datos del Pleistoceno y Holoceno se omiten en ambas figuras. Se observa que los periodos con mayor cantidad de roca sedimentaria tienden a correlacionarse con un mayor número de especies fósiles. Imagen tomada de Foote & Miller (2007).
1.2.2 Potencial de preservación del registro fósil
El potencial de preservación del registro fósil es notablemente bajo y está sujeto a varios sesgos tafonómicos, lo que resulta en una preservación desigual de tejidos mineralizados, partes corporales y grupos taxonómicos. Por ejemplo, es evidente que los grupos que carecen de partes mineralizadas, como los poliquetos, tienen una probabilidad de preservación mucho más baja, con una probabilidad de preservación por género e intervalo de tiempo de solo 0.05. En contraste, los grupos que están compuestos por múltiples partes mineralizadas, como los trilobites, tienen una probabilidad de preservación más alta, que varía entre 0.7 y 0.9 por género e intervalo de tiempo (Tablas 1.1 y 1.2). Esto destaca cómo la composición y las características de los organismos desempeñan un papel crucial en su potencial de preservación en el registro fósil.
| Grupo | Probabilidad |
|---|---|
| Esponjas | 0.4–0.45 |
| Corales | 0.4–0.5 |
| Poliquetos | 0.05 |
| Crustáceos malacostracos | 0.2–0.35 |
| Ostrácodos | 0.5 |
| Trilobites | 0.7–0.9 |
| Briozoos | 0.7–0.75 |
| Braquiópodos | 0.9 |
| Crinoideos | 0.4 |
| Asterozoos | 0.25 |
| Equinoideos | 0.55–0.65 |
| Bivalvos | 0.45–0.5 |
| Gastrópodos | 0.4–0.55 |
| Cephalópodos | 0.8–0.9 |
| Graptólitos | 0.65–0.9 |
| Conodontos | 0.7–0.9 |
| Peces cartilaginosos | 0.1–0.15 |
| Peces óseos | 0.15–0.3 |
| Grupo | Nivel taxonómico | Porcentaje |
|---|---|---|
| Esponjas | Familia | 48 |
| Corales | Familia | 32 |
| Poliquetos | Familia | 35 |
| Crustáceos malacostracos | Familia | 19 |
| Ostrácodos | Familia | 82 |
| Ostrácodos | Género | 42 |
| Briozoos | Familia | 74 |
| Braquiópodos | Familia | 100 |
| Braquiópodos | Género | 77 |
| Crinoideos | Familia | 50 |
| Asterozoos | Familia | 57 |
| Asterozoos | Género | 5 |
| Equinoideos | Familia | 89 |
| Equinoideos | Género | 41 |
| Bivalvos | Familia | 95 |
| Bivalvos | Género | 76 |
| Gastrópodos | Familia | 59 |
| Cephalópodos | Familia | 20 |
| Peces cartilaginosos | Familia | 95 |
| Peces óseos | Familia | 62 |
| Arácnidos | Género | 2 |
El registro fósil, aunque valioso, proporciona una muestra muy limitada de la vida en el pasado. La falta de completitud en este registro ha sido un tema ampliamente discutido en la paleontología. Sin embargo, es importante destacar dos aspectos cruciales:
Diseño de Experimentos Paleontológicos: A pesar de la imperfección del registro fósil, los paleontólogos pueden diseñar experimentos y análisis que tengan en cuenta esta limitación. Esto implica utilizar métodos estadísticos y enfoques que consideren la falta de datos y permitan realizar inferencias basadas en la información disponible.
Sesgo del Registro Fósil: Aunque el registro fósil está lejos de ser completo, el sesgo que presenta puede ser beneficioso desde una perspectiva científica. Cuando los datos recopilados muestran un patrón inesperado o contrario a lo que se podría esperar dada la falta de información, esto puede proporcionar información valiosa sobre la paleobiología.
En resumen, aunque el registro fósil es limitado y puede no proporcionar una imagen completa de la vida en el pasado, los paleontólogos pueden superar esta limitación mediante enfoques estadísticos y considerando el sesgo del registro. Esto permite obtener información fiable y valiosa sobre la evolución y la biología de especies extintas (Fig. 1.47).
Figura 1.47: Comparación de abundancias de especies en una comunidad viva y sus restos subfósiles en un arroyo de marea en California. El gráfico muestra que las especies más abundantes en la muestra viva también tienden a ser más abundantes en la muestra de conchas muertas, con abundancias muertas generalmente más altas que las vivas. Este análisis es clave para comprender la relación entre las comunidades vivas y los fósiles. Imagen tomada de Foote & Miller (2007).
1.2.3 Medidas de la (in)completez del registro fósil
La evaluación de la completitud del registro fósil implica el uso de varias medidas que nos ayudan a comprender cuán bien se representa la historia de la vida en los fósiles. Aquí se detallan tres de estas medidas clave:
- Probabilidad de Muestreo por Unidad de Intervalo de Tiempo (Fig. 1.48): Esta probabilidad se calcula como el número de intervalos temporales en los que esa especie aparece dividido entre todos los intervalos potenciales en los que podría haber aparecido, considerando desde su primera hasta su última aparición en el registro fósil. Por ejemplo, si una especie se registra en 1 de los 4 intervalos temporales potenciales entre su primera y última aparición, su probabilidad de muestreo se calcula como 1 dividido por 4, lo que equivale a 0.25. Es importante señalar que realizar este cálculo para una sola especie puede resultar en un amplio margen de error debido a la variabilidad en los datos. Por esta razón, es común calcular estas probabilidades para conjuntos de especies, lo que proporciona estimaciones más robustas y representativas de la probabilidad de muestreo en el registro fósil.

Figura 1.48: Ilustración esquemática del análisis de intervalos utilizado para estimar la probabilidad promedio de muestreo de un grupo de especies. Cada marca X representa un intervalo de tiempo en el que se muestrea la especie correspondiente; los espacios en blanco intermedios son intervalos en los que no hay registro de la especie. Se tabulan las ocurrencias de cada especie como el número de intervalos en los que se encuentra, excluyendo los de primera y última aparición. Esto se compara con el número de oportunidades de muestreo, es decir, el número de intervalos de tiempo durante los cuales la especie tuvo la posibilidad de ser muestreada o no. Esto se calcula como la suma de los intervalos de tiempo entre la primera y última aparición. Dado que una especie necesariamente debe ser muestreada en sus intervalos de primera y última ocurrencia, estos no se incluyen en las tabulaciones para no sobreestimar la probabilidad de muestreo. En este caso, hay 16 oportunidades de muestreo y 5 ocurrencias, ambos números excluyen las primeras y últimas apariciones. Por lo tanto, la probabilidad de muestreo estimada para estas especies es de 5/16, o 31 por ciento por intervalo de tiempo. Imagen tomada de Foote & Miller (2007).
- Proporción de Taxones Conocidos desde un Solo Intervalo Estratigráfico (Fig. 1.49): Para calcular esta medida, se parte del principio de que un muestreo incompleto tiende a acortar los rangos estratigráficos observados. Por lo tanto, la proporción de taxones conocidos a partir de un solo intervalo estratigráfico se considera inversamente proporcional a la calidad del muestreo. Esto significa que, si la calidad del muestreo es baja, se espera que una mayor proporción de taxones sea conocida solo a partir de un solo intervalo estratigráfico. La fórmula para estimar la probabilidad de muestreo por unidad de tiempo se basa en esta idea y tiene en cuenta las proporciones de especies con rangos estratigráficos de un solo intervalo, dos intervalos y tres intervalos.
Figura 1.49: Distribución de frecuencias de rangos estratigráficos de taxones en dos ejemplos con diferentes niveles de calidad de muestreo, bajo la suposición de un muestreo uniforme. La probabilidad de muestreo por intervalo de tiempo puede estimarse a partir de las frecuencias de taxones con rangos de uno, dos y tres intervalos. Imagen tomada de Foote & Miller (2007).
- Estimación de la Completitud del Registro Fósil a lo Largo del Tiempo: Para evaluar cuán completo es el registro fósil en un período de tiempo extenso, como el Fanerozoico, los paleontólogos hacen estimaciones sobre el número total de especies que se cree que han existido durante ese período y lo comparan con la cantidad de especies conocidas en el registro fósil. Para realizar estas estimaciones, los expertos en paleontología utilizan suposiciones basadas en la longevidad promedio de las especies, que se estima en alrededor de 4 millones de años, y el supuesto de que la diversidad de especies se ha mantenido relativamente constante a lo largo del Fanerozoico. Bajo estas asunciones, se estima que, en promedio, alrededor del 25% de las especies existentes se extinguieron y fueron reemplazadas por nuevas especies cada millón de años. Aplicando esta estimación a lo largo de los 550 millones de años del Fanerozoico, podemos realizar algunos cálculos reveladores. Comenzamos con la diversidad actual, que consiste en aproximadamente 180,000 especies de grupos con un buen registro paleontológico, lo que significa que se han conservado bien como fósiles y han sido bien documentadas por paleontólogos. Luego, consideramos que, en el último medio billón de años, aproximadamente el 25% de la diversidad se reemplazó cada millón de años. Esto implica que durante ese tiempo se originaron alrededor de 25 millones de especies. Sin embargo, cuando comparamos esta estimación de 25 millones de especies con las 300,000 especies fósiles que se han descrito, se hace evidente que solo una pequeña fracción, alrededor del 1% o 2%, de las especies que existieron en el pasado se conocen a través de fósiles. Esta comparación resalta la notable limitación del registro fósil, ya que gran parte de la diversidad de especies que alguna vez poblaron la Tierra permanece en gran medida desconocida para nosotros.
1.2.4 Muestreo del registro fósil
El muestreo del registro fósil es una consideración fundamental en paleontología, y en lugar de preguntarnos si el registro fósil es completo, debemos preguntarnos si es adecuado para el propósito específico que tenemos en mente. Al hacerlo, dos aspectos importantes surgen en el diseño de nuestros estudios paleontológicos:
¿Es una muestra aleatoria? La respuesta es no. Incluso si intentamos realizar un muestreo de manera aleatoria, el registro fósil está sesgado en varios aspectos, como lo hemos discutido previamente. Por lo tanto, debemos ser precavidos al plantear preguntas o resolver problemas específicos utilizando el registro fósil, ya que el sesgo en los datos puede influir en los resultados y conclusiones.
¿Son nuestras medidas sensibles al tamaño muestral? Aquí entra en juego el concepto de “rarefacción”, que es un método utilizado para estimar el número de taxones que habríamos encontrado en una muestra más pequeña. En otras palabras, dado que nuestras muestras del registro fósil pueden variar en tamaño y extensión, la rarefacción nos permite estandarizar y comparar los datos entre diferentes muestras, incluso si son de diferentes tamaños.
La rarefacción es un método utilizado en ecología y paleontología para estandarizar las mediciones de biodiversidad o riqueza taxonómica cuando se trabaja con muestras de diferentes tamaños (Fig. 1.50). La idea principal detrás de la rarefacción es estimar cuántos taxones o especies habrían sido observados en una muestra de un tamaño específico si la muestra más grande se hubiera reducido al tamaño de la muestra más pequeña. En el ámbito de la paleontología, esta técnica se emplea frecuentemente al examinar el registro fósil, donde las dimensiones de las muestras pueden fluctuar debido a la cantidad variable de material fósil disponible. Cuando se aplica la rarefacción, los paleontólogos pueden ajustar los datos procedentes de distintas muestras, lo que simplifica la evaluación de la diversidad y riqueza taxonómica en diferentes conjuntos de fósiles.
Figura 1.50: Ejemplo de curva de rarefacción. Imagen tomada de Foote & Miller (2007).
Los pasos a seguir para llevar a cabo la rarefacción son los siguientes:
Se elige un tamaño de muestra estándar o “tamaño de rarefacción” al que se reducirán todas las muestras para que sean comparables.
Se calcula el número de especies o taxones observados en la muestra original de tamaño completo.
A continuación, se reduce la muestra de tamaño completo al tamaño de rarefacción seleccionado, eliminando aleatoriamente un número específico de especímenes para que la muestra alcance el tamaño deseado, y se calcula el número de especies recogido.
Se repite este proceso varias veces para obtener una estimación promedio de la riqueza taxonómica ajustada al tamaño de la muestra.
Por lo tanto, la curva de rarefacción es una herramienta que muestra cuántas especies se han encontrado en muestras aleatorias de diferentes tamaños. La forma de esta curva es útil para evaluar dos aspectos clave en un yacimiento o formación fósil: (1) si se ha explorado adecuadamente la diversidad de un yacimiento fósil y (2) si es necesario recolectar más muestras para descubrir más especies. Si la curva alcanza una asintota, indica que se ha alcanzado un buen número de muestras y que se tiene una representación adecuada de la diversidad de especies en el yacimiento. Si la curva sigue en aumento y no se estabiliza, sugiere que todavía se pueden encontrar más especies. En este caso, aumentar el esfuerzo de muestreo podría ser beneficioso para descubrir una mayor diversidad. Es importante destacar que este efecto tiende a ser menos pronunciado a niveles taxonómicos más altos debido al anidamiento de las categorías taxonómicas (Fig. 1.51). En cualquier caso, es fundamental tener en cuenta que la curva de rarefacción no puede utilizarse para realizar extrapolaciones más allá de los datos observados, ya que se basa en los resultados actuales de muestreo y no puede predecir la diversidad futura sin una recolección de datos adicional.
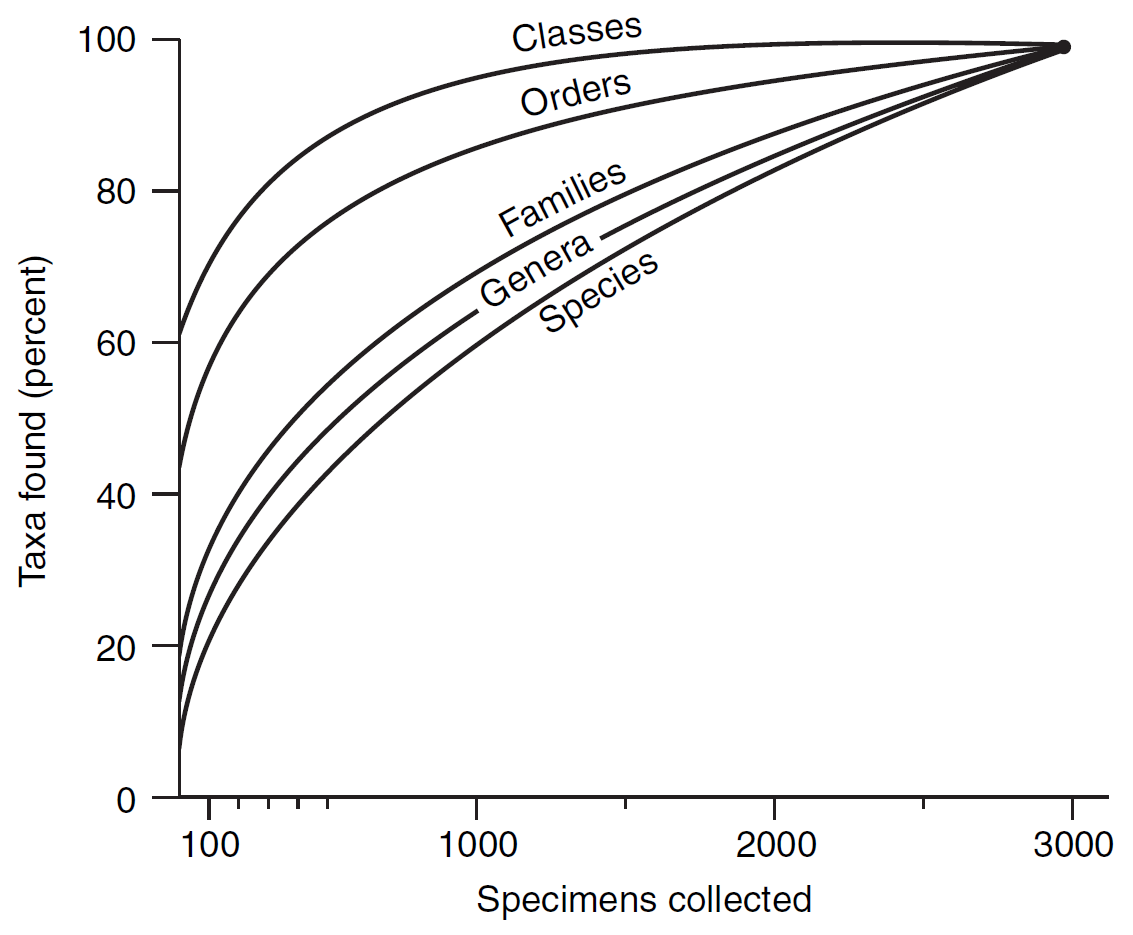
Figura 1.51: Curvas de rarefacción para fósiles de moluscos encontrados en una buena muestra de edad miocena en Dinamarca. El punto en la parte superior derecha representa la muestra real. Las curvas estiman cuántos taxones se habrían encontrado si la muestra hubiera sido más pequeña. Imagen tomada de Foote & Miller (2007).
La curva de rarefacción se calcula siguiendo estos pasos:
Selección de muestras decrecientes en el número de individuos: Se toman muestras del yacimiento o formación fósil en las que el número de individuos disminuye progresivamente. Por ejemplo, comienza con una muestra grande que contiene muchos individuos, como 3000, y luego toma muestras sucesivamente más pequeñas, como 2000, 1000, 500 individuos, y así sucesivamente.
Conteo de especies: Para cada una de estas muestras decrecientes en tamaño, se cuenta cuántas especies diferentes se han encontrado. Esto se hace para cada una de las muestras que se recolectan.
Gráfico de especies vs. número de muestras: Luego, se construye un gráfico que representa el número de especies (en el eje vertical) frente al número de muestras (en el eje horizontal). Cada muestra se representa como un punto en el gráfico. A medida que disminuye el tamaño de las muestras, la cantidad de especies encontradas debería aumentar.
Análisis de la curva: Al observar el gráfico resultante, se puede analizar la forma de la curva de rarefacción.
Ejercicio práctico:
Visualización de la Calidad del Muestreo en Europa a Través de Curvas de Rarefacción
En este ejercicio, nos proponemos visualizar la calidad del muestreo en Europa en dos momentos temporales distintos mediante la construcción de curvas de rarefacción. Se espera que, en el intervalo temporal más reciente, la calidad del muestreo sea mejor, lo que se reflejaría en una curva de rarefacción más asintótica. Esto indica una mayor captura de la diversidad paleobiológica, lo que sugiere un muestreo más exhaustivo y representativo.
Los datos utilizados en este análisis provienen de la Paleobiology Database, y realizaremos todos los análisis utilizando el lenguaje de programación R en el entorno de RStudio.
Antes de empezar, es aconsejable que miremos algunos tutoriales y cursos que nos serán útiles:
Tutorial de descarga e instalación de R y RStudio.
Curso introductiorio a R.
Tutorial de la Paleobiology Database.
Curso introductiorio al uso de la Paleobiology Database.
Descargaremos estos archivos que contienen datos sobre ocurrencias a nivel de género en Europa, correspondientes a los períodos de 750 a 550 ma y 200 a 0 ma. Los datos provienen de la sección de descarga de datos de la Paleobiology Database.
Ejemplo de Submuestreo para Curva de Rarefacción
En este primer ejercicio obtendremos el número de especies que esperamos encontrar en una submuestra de nuestra muestra original. Esto nos dará un punto de datos que forma parte de la curva de rarefacción. Para construir esta curva, debemos repetir el proceso de submuestreo varias veces, tomando diferentes submuestras de tamaños crecientes (por ejemplo, 50, 100, 150, etc.) y anotando cuántas especies únicas encontramos en cada submuestra. Luego, representamos estos datos en un gráfico donde el eje X es el tamaño de la submuestra y el eje Y es el número de especies encontradas.
# Cargamos la muestra de ocurrencias de los géneros de Europa entre 750 y 550 m.a
genus <- read.csv("Data/genus_europe 750 550.csv") # Leer el archivo CSV y asignarlo a "genus"
# Transformamos "genus" en un vector
genus <- as.vector(genus[, 1]) # Extraer la primera columna y convertirla en un vector
# Extraemos el número total de especímenes en la muestra
num_specimens <- length(genus) # Contar el número total de elementos en el vector
print(num_specimens) # Imprimir el número total de especímenes## [1] 328# Extraemos el número total de especies en la muestra
num_species <- length(unique(genus)) # Contar el número de especies únicas
print(num_species) # Imprimir el número total de especies## [1] 106# ¿Cuántos especímenes esperaríamos en una submuestra de 250 especímenes?
# Número de especímenes en la submuestra
n_sub <- 250 # Establecer el tamaño de la submuestra
# Generamos la submuestra (muestreo aleatorio sin reemplazo)
subsample <- sample(genus, n_sub, replace = FALSE) # Realizar el muestreo aleatorio
# Extraemos las especies en la submuestra
unique_species <- unique(subsample) # Obtener las especies únicas en la submuestra
print(unique_species) # Imprimir las especies únicas de la submuestra## [1] "Hiemalora" "Tanarium" "Stratifera" "Multifronsphaeridium"
## [5] "Pteridinium" "Aspidella" "Charnia" "Leiosphaeridia"
## [9] "Dickinsonia" "Yelovichnus" "Ediacaria" "Vaveliksia"
## [13] "Onega" "Tribrachidium" "Palaeopascichnus" "Siphonophycus"
## [17] "Simia" "Vendia" "Kimberella" "Lossinia"
## [21] "Palaeophragmodictya" "Armillifera" "Yorgia" "Tawuia"
## [25] "Kullingia" "Tridia" "Paravendia" "Charniodiscus"
## [29] "Boxonia" "Paliella" "Cyanorus" "Bonata"
## [33] "Phycodes" "Andiva" "Beltanelloides" "Appendisphaera"
## [37] "Parvancorina" "Variomargosphaeridium" "Glaessnerina" "Cyathinema"
## [41] "Synsphaeridium" "Pterospermopsimorpha" "Ovatoscutum" "Cloudina"
## [45] "Rangea" "Chuaria" "Bomakellia" "Densisphaera"
## [49] "Beltanelliformis" "Swartpuntia" "Nimbia" "Cerebrosphaera"
## [53] "Intrites" "Cavaspina" "Coneosphaera" "Beltanella"
## [57] "Archangelia" "Asseserium" "Nemiana" "Anfesta"
## [61] "Temnoxa" "Palaeoplatoda" "Oscillatoriopsis" "Solza"
## [65] "Labruscasphaeridium" "Vendotaenia" "Archaeaspinus" "Spirosolenites"
## [69] "Inaria" "Ventogyrus" "Cochlichnus" "Calyptrina"
## [73] "Protodipleurosoma" "Coniunctiophycus" "Ausia" "Korilophyton"
## [77] "Urasphaera" "Namacalathus" "Pomoria" "Kuckaraukia"
## [81] "Platysolenites" "Medusinites" "Ceratosphaeridium" "Treptichnus"
## [85] "Sinotubulites" "Gyrolithes" "Cambrotubulus" "Skolithos"
## [89] "Palaeolyngbya" "Irridinitus" "Renalcis" "Protolagena"# Contamos el número de especies únicas en la submuestra
num_unique_species <- length(unique_species) # Contar el número de especies únicas
print(num_unique_species) # Imprimir el número de especies únicas en la submuestra## [1] 92Evaluación de la Saturación en la Riqueza de Especies: Construcción de Curvas de Rarefacción
En este ejercicio, evaluaremos el grado de saturación en la riqueza de especies en dos intervalos temporales (750-550 millones de años y 200-0 millones de años) utilizando curvas de rarefacción normalizadas. Para cada intervalo temporal, extraeremos submuestras de tamaños crecientes (0%, 10%, 20%, … 100% del total de especímenes) y registraremos el número de especies únicas en cada una. Los resultados se normalizan dividiendo cada valor entre el máximo número de especies en la muestra completa, permitiéndonos comparar directamente la forma de ambas curvas. El gráfico resultante, con el tamaño relativo de la submuestra en el eje X y el número de especies normalizado en el eje Y, nos permite evaluar visualmente si alguna de las curvas se aproxima a una asíntota más rápidamente que la otra. Esto indicaría que en ese intervalo temporal se ha alcanzado una mayor representatividad en el registro fósil.# Cargamos los datos de ocurrencias de géneros
genus <- read.csv("Data/genus_europe 750 550.csv") # Cargar archivo CSV y asignarlo a la variable "genus"
# Convertimos la columna de géneros en un vector para su análisis
genus <- as.vector(genus[,1]) # Extraer solo la primera columna (géneros) y convertirla en un vector
# Definimos una secuencia de tamaños de muestra para realizar la curva de rarefacción
seq <- seq(0, length(genus), by = length(genus) / 100) # Secuencia que va de 0 al total de especímenes, en 100 pasos
# Inicializamos un vector vacío para almacenar el número de especies únicas en cada submuestra
subsamples <- c()
for (i in 1:length(seq)) {
# Realizamos una submuestra aleatoria sin reemplazo y contamos las especies únicas
subsamples[i] <- length(unique(sample(genus, seq[i], replace = FALSE)))
}
# Graficamos la curva de rarefacción para Europa entre 750 y 550 m.a.
plot(seq, subsamples, main = "Curva de Rarefacción para Europa (750-550 m.a.)",
xlab = "Número de especímenes", ylab = "Número de especies", pch = 19)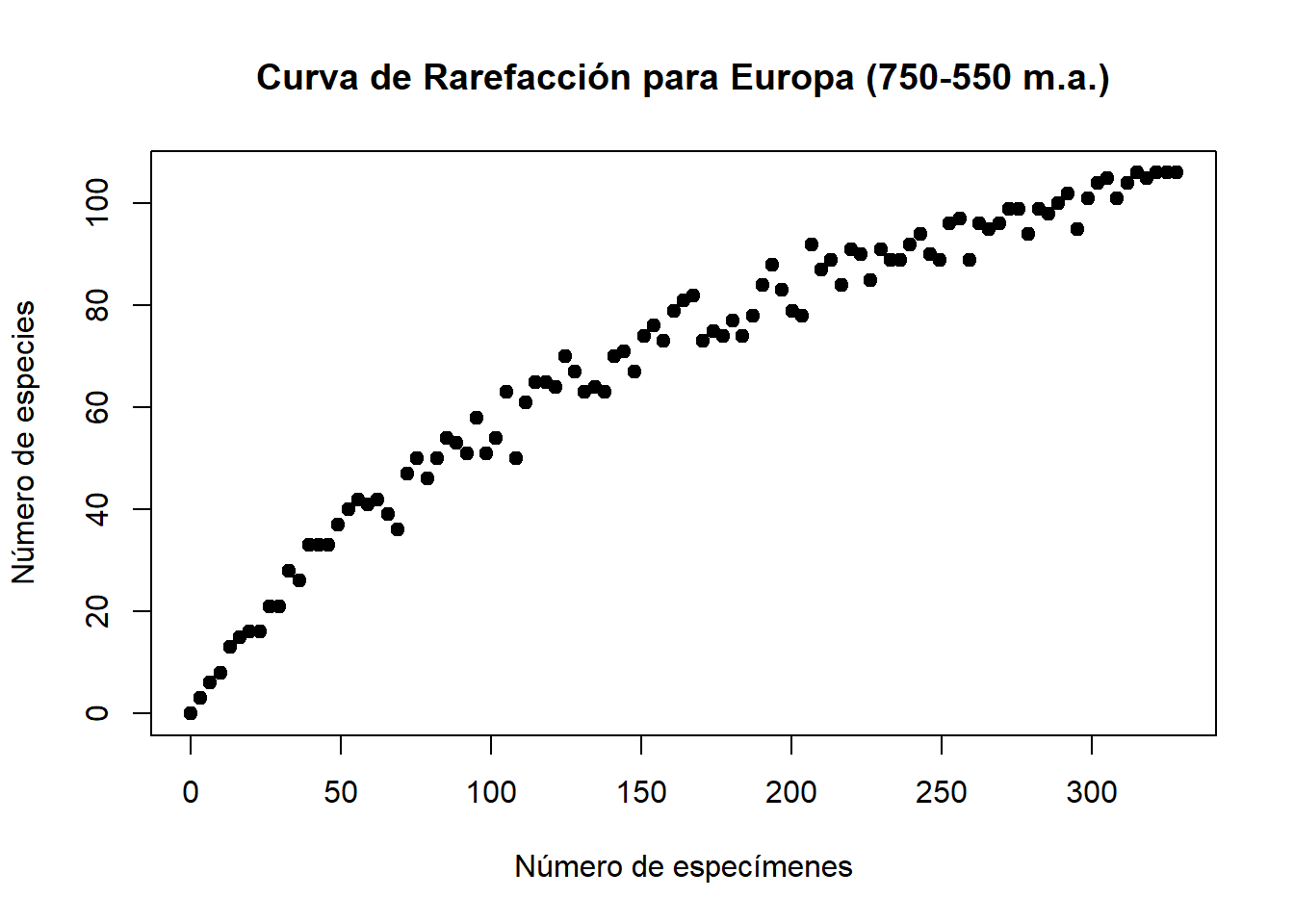
# Repetimos el mismo proceso para Europa entre 200 y 0 m.a.
genus <- read.csv("Data/genus_europe 200 0.csv") # Cargar el archivo de datos de géneros entre 200-0 m.a.
genus <- as.vector(genus[,1]) # Convertir en vector de géneros
seq <- seq(0, length(genus), by = length(genus) / 100) # Crear secuencia de 100 submuestras
subsamples <- c()
for (i in 1:length(seq)) {
# Submuestra aleatoria y cálculo de especies únicas
subsamples[i] <- length(unique(sample(genus, seq[i], replace = FALSE)))
}
# Graficamos la curva de rarefacción para Europa entre 200 y 0 m.a.
plot(seq, subsamples, main = "Curva de Rarefacción para Europa (200-0 m.a.)",
xlab = "Número de especímenes", ylab = "Número de especies", pch = 19)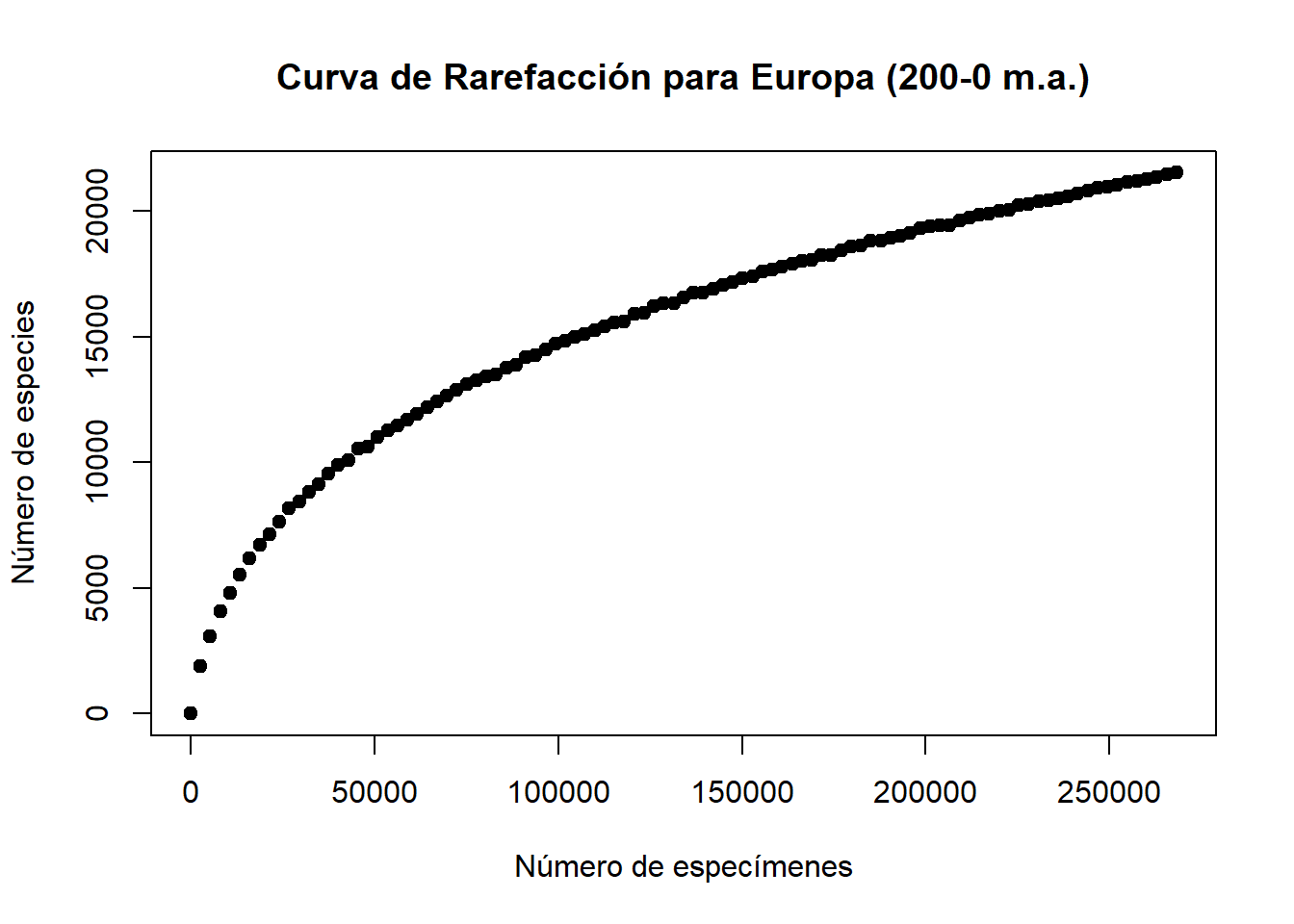
# Comparación de ambas curvas de rarefacción en una sola gráfica
# Datos para Europa entre 750 y 550 m.a.
genus1 <- read.csv("Data/genus_europe 750 550.csv")
genus1 <- as.vector(genus1[,1])
seq1 <- seq(0, length(genus1), by = length(genus1) / 100)
subsamples1 <- c()
for (i in 1:length(seq1)) {
subsamples1[i] <- length(unique(sample(genus1, seq1[i], replace = FALSE)))
}
# Datos para Europa entre 200 y 0 m.a.
genus2 <- read.csv("Data/genus_europe 200 0.csv")
genus2 <- as.vector(genus2[,1])
seq2 <- seq(0, length(genus2), by = length(genus2) / 100)
subsamples2 <- c()
for (i in 1:length(seq2)) {
subsamples2[i] <- length(unique(sample(genus2, seq2[i], replace = FALSE)))
}
# Creación del dataframe para graficar ambas curvas
df <- data.frame(
Numero_de_Especimenes = c(1:101,1:101),
Numero_de_Especies = c(subsamples1 / max(subsamples1), subsamples2 / max(subsamples2)),
Periodo = as.factor(c(rep("750-550 m.a.", length(seq1)), rep("200-0 m.a.", length(seq2))))
)
# Graficamos ambas curvas de rarefacción en la misma gráfica con plot()
# Separamos los datos por período
df_750_550 <- subset(df, Periodo == "750-550 m.a.")
df_200_0 <- subset(df, Periodo == "200-0 m.a.")
# Establecemos los límites del gráfico basados en el rango de datos
plot(range(df$Numero_de_Especimenes), range(df$Numero_de_Especies), type = "n", # Gráfico vacío para establecer los límites
xlab = "Número de especímenes (% sobre el total)",
ylab = "Número de especies (normalizado)",
main = "Comparación de Curvas de Rarefacción en Europa")
# Añadimos los puntos para cada período en diferentes colores
points(df_750_550$Numero_de_Especimenes, df_750_550$Numero_de_Especies, col = "black", pch = 19)
points(df_200_0$Numero_de_Especimenes, df_200_0$Numero_de_Especies, col = "grey", pch = 19)
# Opcionalmente, añadimos una leyenda para mayor claridad
legend("bottomright", legend = c("750-550 m.a.", "200-0 m.a."), col = c("black", "grey"), pch = 19)
1.3 La especie en paleontología
1.3.1 El concepto de especie
El concepto de especie es fundamental en biología y existen varios enfoques para definir lo que constituye una especie.
Concepto de Especie Biológica (según Dobzhansky y Mayr): Según este enfoque, una especie es una serie de poblaciones que pueden cruzarse de manera real o potencial para producir descendientes viables y están reproductivamente aisladas de otras poblaciones en condiciones naturales. Este concepto se basa en la capacidad de las poblaciones para intercambiar genes a través de la reproducción sexual y, por lo tanto, se aplica principalmente a organismos que se reproducen sexualmente.
El concepto de especie biológica implica la existencia de barreras reproductivas que pueden ser de naturaleza geográfica o biológica, lo que resulta en la separación de linajes evolutivos. Esta perspectiva considera a las especies como unidades genéticas y evolutivas independientes, libres de seguir su propio curso en respuesta a procesos genéticos y cambios ambientales. Sin embargo, este concepto tiene limitaciones:
Inaplicabilidad en el Registro Fósil: No se puede aplicar el concepto de especie biológica a organismos extintos, ya que no es posible observar directamente su capacidad de reproducción.
Limitaciones en Organismos Asexuales: Este enfoque es limitado en organismos que se reproducen asexualmente, ya que no involucran la reproducción sexual ni las barreras reproductivas.
Hibridación: El concepto de especie biológica no aborda adecuadamente casos de hibridación, donde se producen descendientes fértiles a través del cruce de poblaciones que, en teoría, deberían ser reproductivamente aisladas. Esto es común en plantas vasculares y, en algunos casos, en animales.
Concepto de Especie Ecológica: Según este enfoque, una especie se define como un linaje (o conjunto de linajes cercanamente relacionados) que ocupa una zona adaptativa mínimamente diferente en su distribución en comparación con otras especies. Además, se desarrolla independientemente de todos los linajes establecidos fuera de su área biogeográfica de distribución. Este concepto incorpora nociones de nicho ecológico y exclusión competitiva para explicar las diferencias entre especies basadas en su explotación de recursos ecológicos y adaptación a ambientes específicos.
Concepto de Especie Evolutiva: Este concepto se basa en linajes únicos de poblaciones ancestrales y descendientes que evolucionan independientemente de otros grupos similares. A diferencia del concepto biológico, se centra en el aislamiento genético actual en lugar del potencial. Además, considera que la evolución cladogenética puede ser reticulada, lo que significa que poblaciones previamente separadas pueden reunirse nuevamente, dando lugar a especies híbridas.
Concepto de Especie Filogenética: Según este enfoque, una especie es el grupo monofilético más pequeño con una ascendencia común. Esto se basa en la identificación de un carácter derivado o apomórfico compartido por todos los organismos del grupo. Este enfoque tiene en cuenta similitudes genéticas y moleculares para discernir especies.
Concepto de Especie Morfológica: En este concepto, las especies se definen por diferencias anatómicas medibles. Es especialmente útil en paleontología, donde no siempre se pueden evaluar características de reproducción o genéticas. Sin embargo, este enfoque puede ser limitado, ya que no siempre refleja las diferencias en comportamiento o ecología.
Estos enfoques ofrecen diferentes perspectivas sobre cómo definir y reconocer las especies. En algunos casos, pueden aplicarse a ciertos grupos de organismos y en otros no. La diversidad de conceptos de especies refleja la complejidad de la vida y la necesidad de abordar esta complejidad desde múltiples ángulos para comprender mejor la biodiversidad y la evolución.
Las especies suelen presentar una variabilidad interna, lo que se conoce como politipismo. Este término se utiliza en zoología y botánica para describir grupos taxonómicos que muestran más de un subgrupo en el nivel taxonómico más bajo. En otras palabras, se refiere a la existencia de múltiples formas dentro de una especie, que pueden incluir subespecies y variedades. Dentro de una especie, las subespecies se caracterizan por mantener una uniformidad de tipo, es decir, comparten características distintivas en su apariencia o genética. Además, estas subespecies a menudo ocupan áreas geográficas extensas que son ambientalmente uniformes. Por otro lado, algunas variaciones dentro de una especie pueden estar relacionadas con gradientes ambientales. En lugar de subgrupos claramente definidos, la variación clinal implica que las características de una población cambian gradualmente a medida que te alejas de una ubicación geográfica o entorno específico. Esto refleja la adaptación de la especie a diferentes condiciones ambientales a lo largo de su rango. El politipismo y la variación clinal son fenómenos comunes en la naturaleza y subrayan la adaptabilidad de las especies a una variedad de entornos y condiciones. Estos conceptos son esenciales para comprender la diversidad y la evolución en la biología.
Lectura recomendada
Aldhebiani, A. Y. (2018). Species concept and speciation. Saudi journal of biological sciences, 25(3), 437-440.
De Queiroz, K. (2007). Species concepts and species delimitation. Systematic biology, 56(6), 879-886.
1.3.2 Especies y especiación
Los mecanismos de aislamiento reproductivo son fundamentales en la especiación, el proceso mediante el cual una población de organismos se divide en dos o más poblaciones reproductivamente aisladas que evolucionan en direcciones diferentes. Estos mecanismos evolutivos evitan que los individuos de diferentes especies se crucen, lo que es esencial para mantener la integridad genética de las especies.
Aislamiento precigótico: Estos mecanismos actúan antes de la fertilización y previenen la formación de híbridos. Son especialmente importantes porque evitan que los recursos y la energía se desperdicien en la producción de descendencia que podría ser inviable o estéril. Los mecanismos de aislamiento precigótico incluyen:
Aislamiento temporal o del hábitat: Este mecanismo opera cuando las poblaciones ocupan diferentes hábitats, lo que impide que los individuos de diferentes especies se encuentren. También puede ser el resultado de barreras físicas, como ríos o montañas. Incluso diferencias en el tiempo de madurez sexual o floración pueden evitar el apareamiento.
Aislamiento conductual: Las especies animales a menudo tienen rituales de apareamiento específicos que son incompatibles entre especies. Estos rituales incluyen comportamientos específicos, señales visuales o auditivas que solo son efectivos dentro de la misma especie.
Aislamiento mecánico: La estructura de los órganos reproductores de las especies puede ser incompatible. Por ejemplo, los genitales de algunos insectos actúan como “cerraduras” y “llaves”, y solo permiten el apareamiento con individuos de la misma especie que tengan estructuras complementarias.
Aislamiento gamético: Incluso si los individuos de diferentes especies se encuentran y copulan, en algunos casos, los óvulos y espermatozoides no pueden fusionarse debido a incompatibilidades gaméticas. En algunos entornos, como el desove sincrónico de corales en los arrecifes, varias especies liberan gametos simultáneamente, lo que facilita la hibridación entre especies. Aproximadamente un tercio de los cruces entre especies de coral resultan en híbridos viables, mientras que los dos tercios restantes son incompatibles debido a incompatibilidades de gametos. Por ejemplo, en erizos de mar del género Strongylocentrotus, la concentración de espermatozoides necesaria para fecundar completamente los óvulos de su propia especie solo fertiliza un 1.5% de los óvulos de otras especies. Esta incompatibilidad gamética es común en invertebrados marinos, aunque las causas fisiológicas no son del todo claras. En algunos insectos, como la Drosophila, se ha observado que, tras la inseminación, la hembra experimenta una hinchazón vaginal que impide la fertilización por parte de esperma de otra especie. En plantas, los granos de polen de una especie pueden germinar en el estigma de otra, pero el crecimiento de los tubos polínicos se detiene antes de alcanzar los óvulos, evitando así la fecundación. Este mecanismo, conocido como incongruencia o incompatibilidad cruzada, es común en angiospermas. Además, existe una relación con la autoincompatibilidad: los cruces entre individuos autocompatibles (SC) e individuos autoincompatibles (SI) pueden generar híbridos, pero los cruces recíprocos no, ya que los tubos polínicos no llegan a los óvulos, fenómeno conocido como incompatibilidad unilateral.
Aislamiento postcigótico: Estos mecanismos operan después de la fertilización y evitan el éxito del cruce entre poblaciones. Pueden resultar en la muerte de los híbridos o en su esterilidad. Algunos de estos mecanismos incluyen:
Mortalidad de cigotos y no viabilidad de híbridos: En algunos cruces entre especies, aunque el óvulo es fertilizado, el cigoto resultante no se desarrolla correctamente, lo que provoca la inviabilidad del híbrido y su muerte. Este fenómeno se observa tanto en plantas como en animales. En el género Rana, por ejemplo, la viabilidad del embrión híbrido varía: algunos cigotos no segmentan, otros no completan la gastrulación, y otros muestran fallos en etapas avanzadas del desarrollo. Esto sugiere una diferenciación en los genes de desarrollo entre las especies parentales que impide el desarrollo adecuado del híbrido. De manera similar, en cruces recíprocos de mosquitos del género Culex, las incompatibilidades se deben a interacciones entre genes nucleares y citoplasmáticos, donde el citoplasma heredado de la hembra afecta el desarrollo del híbrido. En las angiospermas, la viabilidad del embrión depende de un endospermo funcional (tejido especializado que se forma en las semillas de las plantas con flores y que proporciona nutrientes al embrión en desarrollo). En cruces interploidales y en algunos cruces entre especies con el mismo nivel de ploidía, el desarrollo del endospermo puede fallar, lo que lleva al aborto del embrión híbrido. Este es uno de los mecanismos de aislamiento reproductivo postcigótico más comunes en plantas.
Esterilidad híbrida: En otros casos, los híbridos pueden ser viables pero estériles. En todos estos casos la esterilidad se debe a los desequilibrios cromosómicos debido al diferente número de cromosomas en las especies parentales Un ejemplo bien conocido de esto es la mula, un híbrido entre un caballo y una burra.
Los tipos principales de especiación según el origen geográfico de las barreras reproductivas son (Fig. 1.52):
Especiación Alopátrica: La especiación alopátrica ocurre cuando una especie se divide en dos grupos que quedan geográficamente aislados, impidiendo que se reproduzcan entre sí. Este aislamiento puede deberse a barreras físicas como montañas, ríos o cuerpos de agua que bloquean el contacto entre las poblaciones. Con el tiempo, cada grupo se adapta a las características y demandas únicas de su entorno, lo que lleva a la acumulación de diferencias genéticas y, eventualmente, a la formación de nuevas especies. Un ejemplo clásico es el de los pinzones de las islas Galápagos, que han desarrollado diferentes formas de pico adaptadas a los alimentos disponibles en cada isla, separadas por el océano. Otro caso interesante es el de las ardillas del Gran Cañón en Arizona: cuando se formó este profundo desfiladero, las poblaciones de ardillas quedaron divididas a ambos lados, incapaces de cruzarlo y reproducirse entre sí. Con el tiempo, se originaron dos especies distintas de ardillas en los bordes norte y sur del cañón. En contraste, aves y otras especies que podían cruzar la barrera continuaron reproduciéndose y no experimentaron este tipo de especiación.
Especiación Peripátrica: La especiación peripátrica ocurre cuando un pequeño grupo de individuos se separa de la población principal, quedando aislado en una región geográfica limitada. Como en la especiación alopátrica, las barreras físicas impiden que los individuos de ambos grupos se reproduzcan entre sí. Sin embargo, en la especiación peripátrica, el tamaño reducido de la población aislada hace que la deriva genética pueda tener un impacto considerable. Esto significa que, debido al tamaño pequeño de esta subpoblación, ciertas características únicas pueden fijarse por puro azar más fácilmente a través de generaciones, acelerando la diferenciación y dando lugar a una nueva especie. Este tipo de especiación suele ocurrir en islas o hábitats aislados donde las pequeñas poblaciones desarrollan rasgos distintivos.
Especiación Parapátrica: La especiación parapátrica ocurre cuando una especie ocupa un área geográfica extensa en la que los individuos no están completamente aislados, pero tienden a reproducirse solo con aquellos de su región inmediata. A diferencia de la especiación alopátrica o peripátrica, donde existen barreras físicas claras, en la especiación parapátrica las poblaciones están separadas por diferencias en el ambiente mismo, lo que genera variaciones en las presiones selectivas de cada área. Un ejemplo es el caso de plantas que crecen en suelos contaminados. Las actividades mineras pueden dejar residuos ricos en metales pesados, como plomo y zinc, que impiden el crecimiento de la mayoría de las plantas. Sin embargo, algunas especies, como el bisonte o césped de búfalo (Bouteloua dactyloides), han desarrollado tolerancia a estos metales y se han adaptado para crecer en estos suelos.
Especiación Simpátrica: La especiación simpátrica ocurre sin una barrera geográfica evidente. En este caso, las poblaciones comparten el mismo espacio, pero desarrollan barreras reproductivas debido a preferencias de hábitat o recursos. Un ejemplo es el mosca de la manzana (Rhagoletis pomonella), que ha comenzado a especializarse en poner huevos en manzanas en lugar de su planta original, el espino, lo que podría llevar a la formación de nuevas especies. Es un concepto controvertido.
Especiación Artificial: La especiación artificial se refiere a la creación de nuevas especies por intervención humana, a menudo a través de experimentos de laboratorio, como en el caso de estudios con drosófilas. Estos experimentos pueden inducir características genéticas específicas que resultan en nuevas especies.
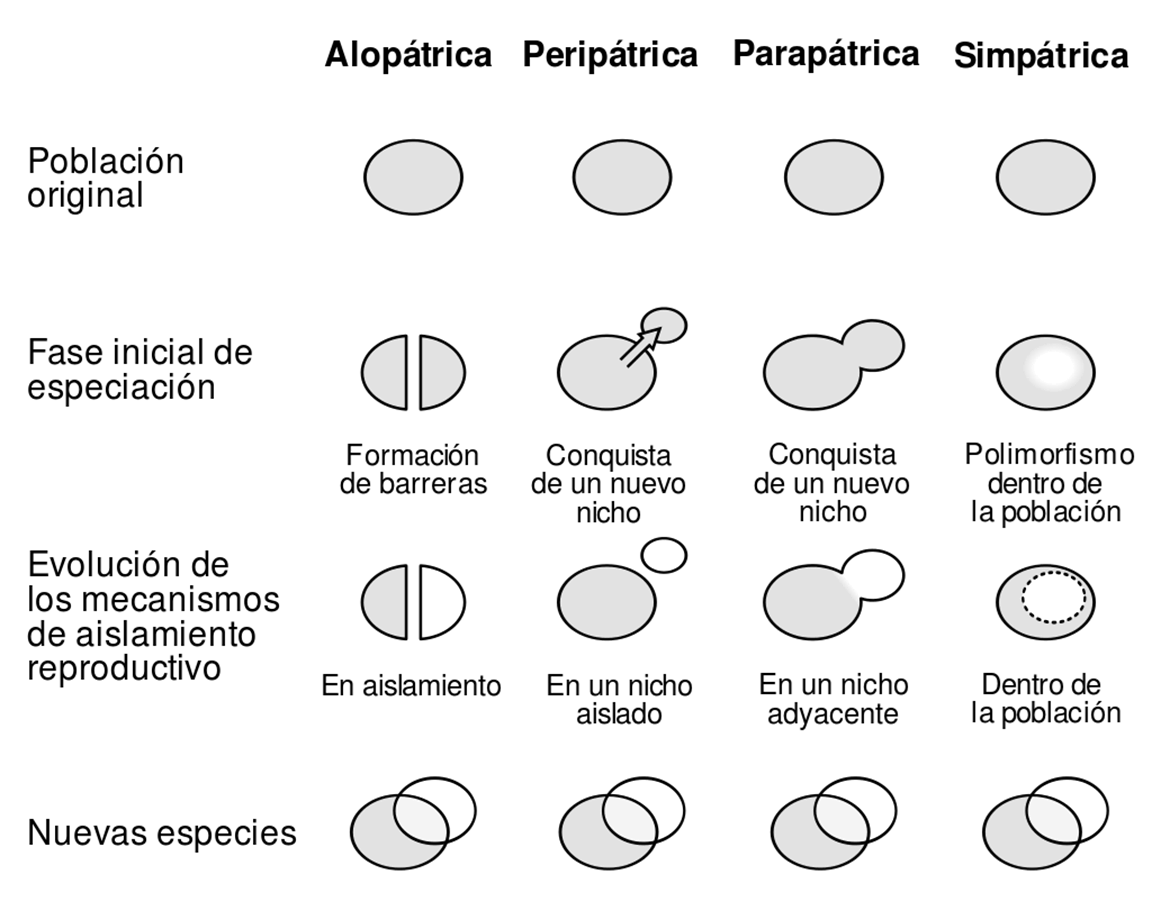
Figura 1.52: Tipos principales de especiación según el origen geográfico de las barreras reproductivas. Imagen disponible bajo licencia Creative Commons CC0.
1.3.3 Reconocimiento de (morfo)especies en el registro fósil
El reconocimiento de especies en el registro fósil es esencial en paleontología y se basa principalmente en criterios morfológicos. Aunque en la práctica, los zoólogos y botánicos a menudo utilizan evidencia genética para apoyar sus clasificaciones, en muchos fósiles, esta evidencia genética está ausente. Por lo tanto, la taxonomía en paleontología se basa en características morfológicas observables, lo cual puede ser desafiante debido a la naturaleza fragmentaria y a menudo parcial de los fósiles.
Sin embargo, a pesar de estas limitaciones, la discriminación de especies en el registro fósil es posible dentro de un margen de confianza. Esto se debe a la alta correlación que existe entre las características morfológicas de los organismos y otros aspectos esenciales de su biología. De hecho una gran cantidad de trabajos muestra una buena correlación entre la distancia morfológica y genética entre pares de especies, tanto en animales como plantas (Fig. 1.53).
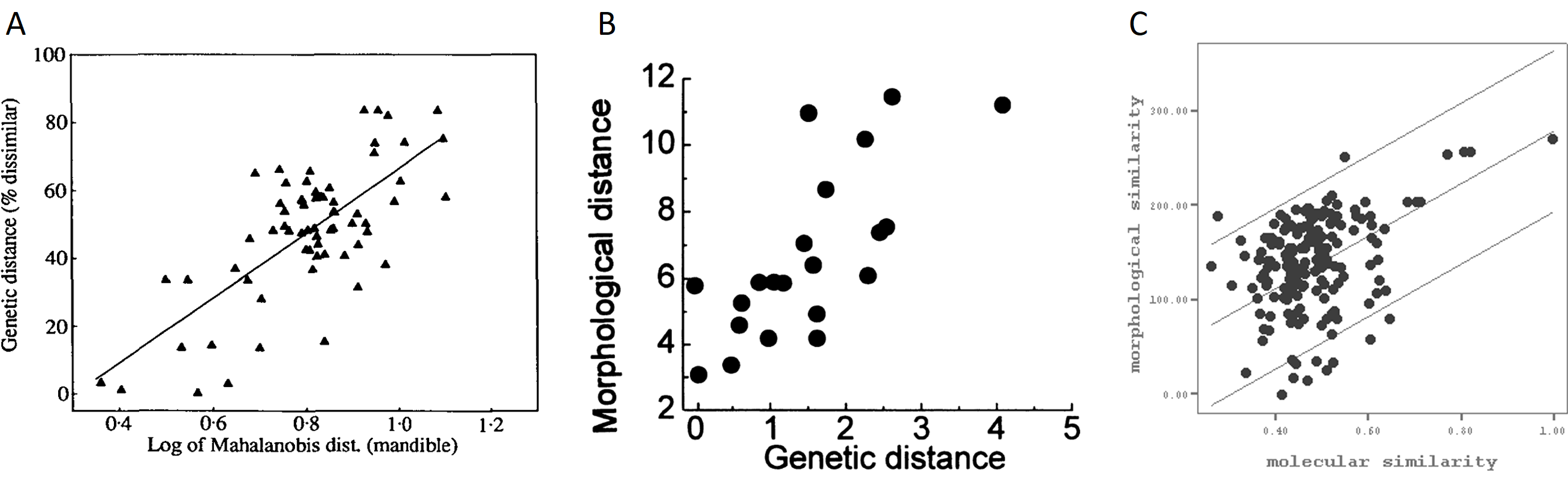
Figura 1.53: Ejemplos de correlación entre distancia morfológica y genética en (A) vertebrados, (B) invertebrados y (C) plantas. Imágenes tomadas de Festing & Roderick (1989), Jackson & Cheetham (1994) y Babić et al. (2012), respectivamente.
En cualquier caso, es obvio que existen ciertas limitaciones en la discriminación de especies en el registro fósil a través de caracteres morfológicos que los paleontólogos deben tener presente:
Caracteres Morfológicos Perdidos: Con el paso del tiempo, es posible que ciertos caracteres morfológicos se pierdan o no se conserven adecuadamente en los fósiles. Esto puede deberse a la degradación de los tejidos blandos o a la erosión de partes del esqueleto. La ausencia de características cruciales puede hacer que la identificación de especies sea más desafiante.
Deformaciones Tafonómicas Sobre la Morfología del Fósil: Durante el proceso de fosilización los fósiles pueden sufrir deformaciones o distorsiones. Esto puede hacer que las características morfológicas de un fósil se vean alteradas, lo que dificulta su correcta identificación y clasificación.
Dimorfismo Sexual: En algunas especies, existe dimorfismo sexual, lo que significa que los individuos de diferentes sexos presentan características morfológicas distintas. En el registro fósil, este fenómeno podría llevar a la identificación errónea de dos especies distintas o a la falta de reconocimiento de una especie dimórfica.
Especies Crípticas: Algunas especies presentan una gran similitud morfológica, lo que dificulta su distinción a través de caracteres visibles. Estas especies crípticas son difíciles de diferenciar solo por sus características morfológicas, lo que puede llevar a una subestimación de la diversidad real en el registro fósil.
Especies Polimórficas: Las especies polimórficas exhiben una variabilidad morfológica entre sus individuos. Esto puede deberse a factores genéticos o ambientales. La variabilidad en la morfología de una especie polimórfica puede llevar a la identificación errónea de diferentes fósiles como especies separadas.
Presencia de Cronoespecies: Las cronoespecies son especies que existieron durante un período de tiempo prolongado y experimentaron cambios morfológicos significativos a lo largo de su historia. Esto puede llevar a la confusión, ya que los fósiles de diferentes etapas de una cronoespecie podrían parecer especies diferentes debido a sus diferencias morfológicas, a pesar de ser parte de la misma especie en evolución.
1.3.4 (Morfo)especies en el registro fósil y tiempo geológico
1.3.4.1 Linajes evolutivos
La evolución es un proceso continuo en el que las poblaciones experimentan cambios a lo largo del tiempo debido a factores como la selección natural, la deriva genética y otros mecanismos. En algunos casos, estos cambios son graduales y no generan barreras reproductivas significativas. Sin embargo, en otros, las transformaciones evolutivas pueden ser tan marcadas que impidan o dificulten la hibridación efectiva entre poblaciones de diferentes períodos geológicos.
La presencia de diferencias morfológicas o de registros fósiles notables entre poblaciones en intervalos de tiempo distintos puede indicar que se están tratando con linajes evolutivos separados. Estos linajes pueden clasificarse como especies o subespecies, dependiendo de la magnitud de las diferencias y de la existencia de barreras reproductivas potenciales.
Este enfoque, que considera los linajes en lugar de ceñirse a una única especie, es particularmente relevante al trabajar con el registro fósil, donde la información genética suele ser escasa. Al abordar la evolución en términos de linajes en lugar de limitarse a las especies, los paleontólogos pueden reflejar de manera más precisa la diversidad y la dinámica evolutiva a lo largo del tiempo geológico.
Normalmente, las especies se pueden reconocer durante largos intervalos de tiempo, mostrando variaciones dentro de un rango morfológico estrecho. Sin embargo, surge la pregunta: ¿podrían hibridar la población inicial y la final? Probablemente no, lo que sugiere que no deberíamos considerar estos conjuntos temporales como una única especie. En cambio, es más apropiado hablar de linajes (Fig. 1.54).
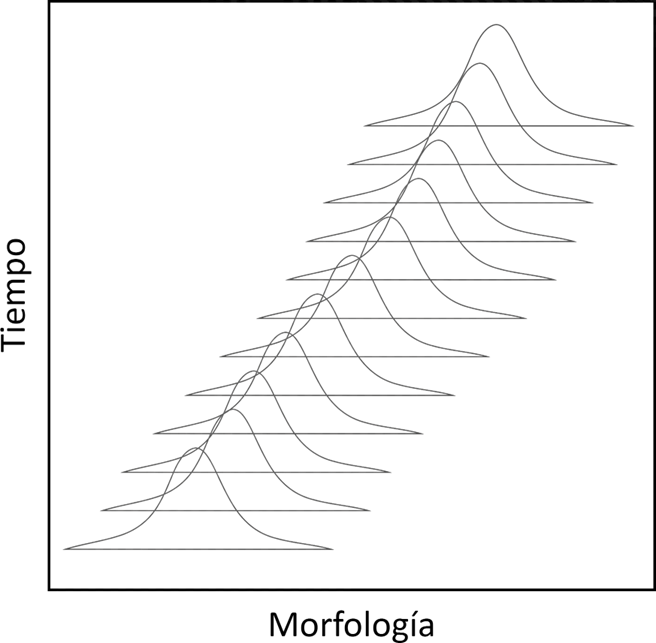
Figura 1.54: Ejemplo gráfico de cómo se documenta el cambio morfológico continuo en una misma población a lo largo del tiempo en el registro fósil.
1.3.4.2 Plasticidad fenotípica y registro fósil
Es importante destacar que la plasticidad fenotípica, es decir, la capacidad de un organismo para mostrar diferentes rasgos en respuesta a cambios en su entorno, puede llevar a interpretaciones erróneas de cambios morfológicos evolutivos en especies extintas. Esto es particularmente relevante cuando se observa variabilidad coetánea en el registro fósil, lo que significa que diferentes individuos de una misma especie extinta pueden mostrar variaciones morfológicas en un período de tiempo específico.
La variabilidad coetánea puede deberse a la plasticidad fenotípica de una especie, lo que significa que los individuos pueden haber desarrollado diferentes características morfológicas en respuesta a condiciones ambientales cambiantes, pero no necesariamente representa un cambio evolutivo. En otras palabras, los cambios observados en la morfología de los fósiles pueden deberse a una respuesta al su entorno en lugar de cambios genéticos a lo largo de generaciones sucesivas (Fig. 1.55).
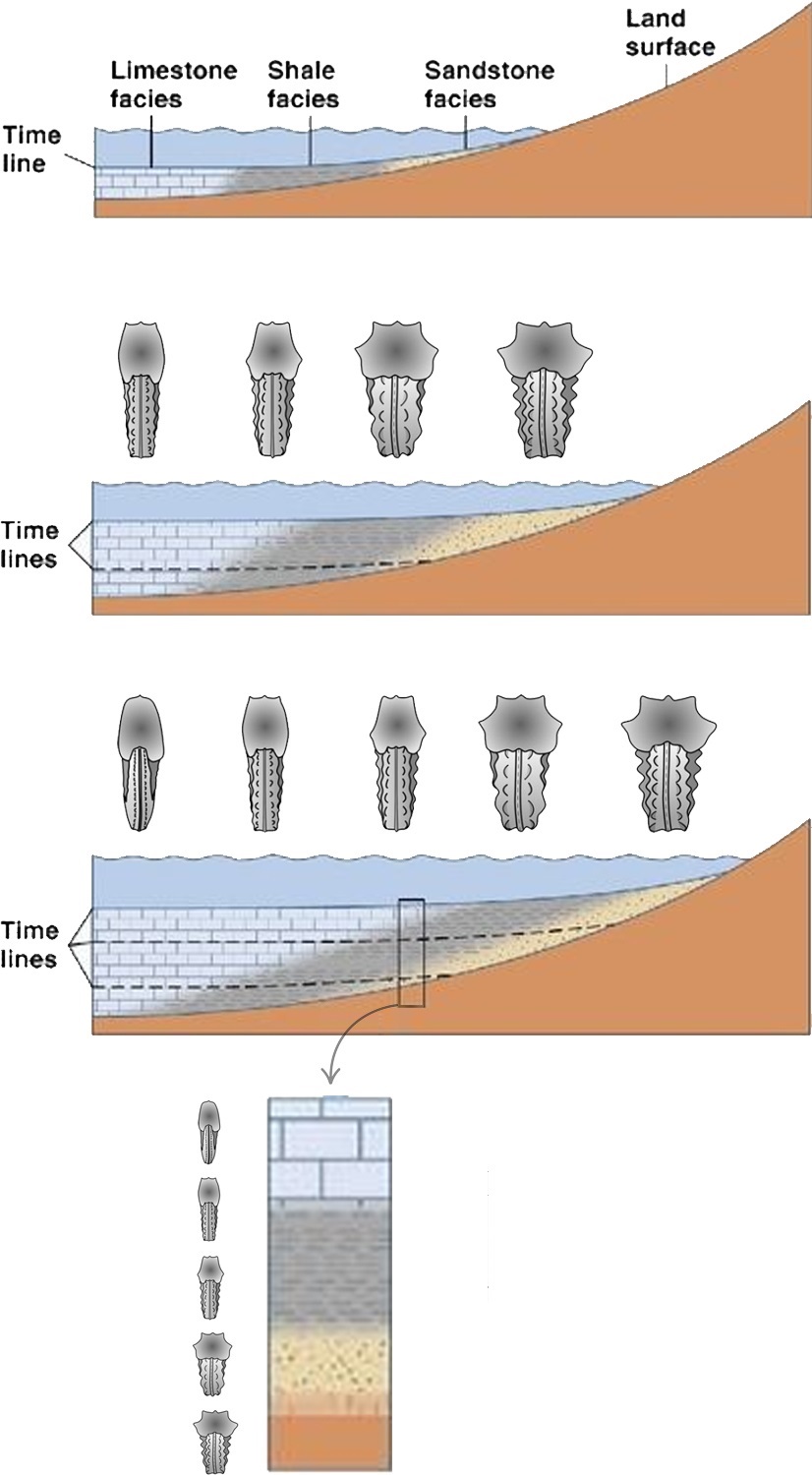
Figura 1.55: Ilustración de un corte transversal de una costa que muestra un proceso de transgresión marina en dirección descendente. En este entorno, las aguas están habitadas por un molusco hipotético que presenta plasticidad fenotípica en su exoesqueleto, variando su morfología en función de la proximidad a la costa. El diagrama inferior representa una columna estratigráfica, en la cual se observa un cambio morfológico a lo largo del tiempo, atribuido exclusivamente a la plasticidad fenotípica de la especie. Imagen modificada de © Geology Desk y Wilmsen & Mosavinia (2011).
Una manera de descartar cambios evolutivos y atribuir la variabilidad morfológica a la plasticidad fenotípica es buscar correlaciones entre los diferentes morfotipos y las facies o los tipos de entorno en los que se encuentran los fósiles. Esta correlacion puede buscarse entre niveles sincronicos o a lo alrgo de series temporales (Fig. 1.56). Si se encuentra dicha correlación entre las características morfológicas y los entornos específicos, esto puede sugerir que la variabilidad morfológica es el resultado de plasticidad fenotípica. Sin embargo, no siempre es posible establecer esta correlación, y en tales casos, la distinción entre plasticidad fenotípica y cambio evolutivo puede ser más desafiante. Por lo tanto, los paleontólogos deben utilizar múltiples líneas de evidencia y ser conscientes de la plasticidad fenotípica al interpretar cambios morfológicos en especies extintas.
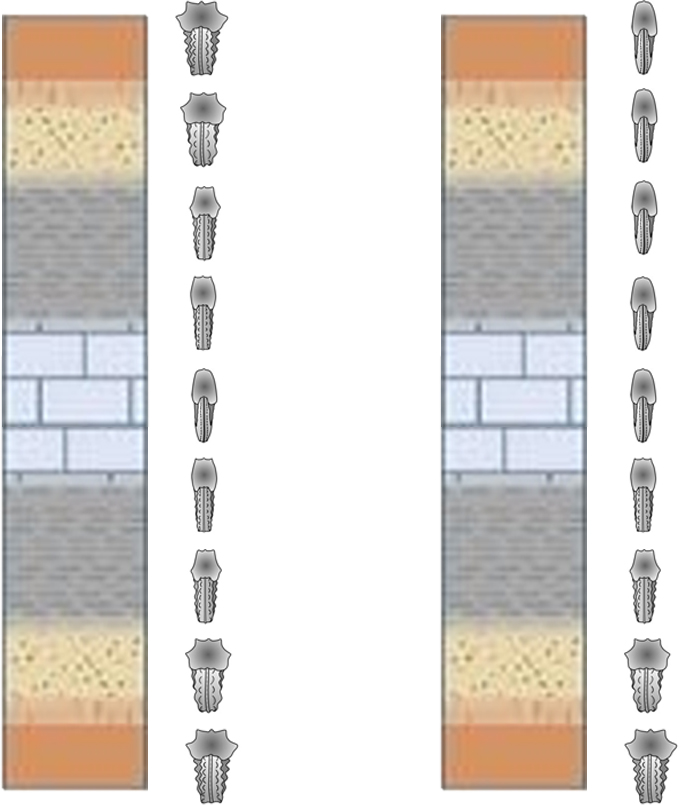
Figura 1.56: Ilustración de dos columnas estratigráficas donde, la de la izquierda muestra una clara correspondencia entre facies y morfotipos, lo que sugiere que los cambios morfológicos en este caso pueden atribuirse a la plasticidad fenotípica. En contraste, la columna de la derecha no refleja tal relación, indicando cambio evolutivo. Imagen modificada de © Geology Desk y Wilmsen & Mosavinia (2011).
Lectura recomendada
Lister, A. M. (2021). Phenotypic plasticity in the fossil record. In Phenotypic Plasticity & Evolution (pp. 267-297). CRC Press.