Chapter 3 Biology of Coral Bleaching
Putnam lab copy of Coral Bleaching: Patterns, Processes, Causes, and Consequences (van Oppen and Lough 2018). This link will only be accessible to Putnam Lab members.
Thermal Response of Algal Symbiont
Chapter 8.2 in the book above
Symbiosis = dependent on productivity of intracellular algal symbionts and requires photosynthetic production in each host cell.
Oxidative theory of coral bleaching:
- ROS generation by algal photosynthesis then overwhelms host and symbiont mechanisms to detoxify ROS and repair oxidative damage.
- High temps and irradiance = large quantify and rate of energy entering/being processed by photosystems (light-harvesting complexes, photosynthetic ETS in algal chloroplast)
- surrounding host membranes and tissues constrain efflux of photosynthetically generated O2 away from algae and import of dissolved inorganic carbon to maintain photosynthetic carbon fixation
- thus susceptible to oxidative damage
- failure of the above is likely driving coral bleaching cascades
Mechanisms of agal thermal stress:

mech
Carotenoid peridinin:
- possesses a higher absorbance of green to blue light relative to chlorophyll.
- symbionts use this together with chlorophyll a and c pigmentation.
- gives the symbionts the golden brown color and dark tone to the coral
- integrated into intrinsic membrane-bound antenna chlorophyll a and c2-peridinin-protein-complex that transfers E to photosystems.

carto
Light goes through the above complexes and enters the ETS from PSII to PSI to ferredoxin-nicotinamide adenine dinucleotide phosphate (NADP+) reductase. Efficiency of this can be assessed by active chlorophyll induction fluorometry.
- Maximum quantum yield of PSII chl-a flourescence = Fv/Fm = proportion of absorbed light E that enters ETS from PSII
- Decline in above = nonspecific photoinactivation due to PSII damage
- reaction center protein D1 = particularly susceptible to photodamage
- PSII and PSI are right next to each other within the thylakoid = bidirectional E transfer can happen, esp during thermal stress
- Because of the above, excess light E can be directed away from PSII to PSI to be converted to heat and quenched by reduced reaction center chlorophyll P700+.
PSII degradation:
- reduces capacity of photosynthetic apparatus to use light E and therefore increased pressure on the remaining PSII units
High O2 env (hyperoxic) = increase in rate of spontaneous/catalyzed ROS production by ETS. Release of ROS from algae is b/c of overexcitation of photosynthetic apparatus.
- mostly occurs via water-water cycle/Mehler rxn: direct reduction of O2 by PSI to superoxide (O2-), which is caused by high light or combination of high light and temperature.
- results in enhanced ATP production at expense of NADPH (antioxidant systems need NADPH)
High light = condensation of thylakoid lamellae
High temp = thylakoid membrane disorganization
Leakage of ROS from algal cell initiates bleaching cascade.
Nitric oxide (NO) = important cellular signaling molecule that is involved in immunity, response to endosymbiotic microbes, and apoptosis pathways
- in high temps and bleaching, symbionts increase NO production
- NO is lipophilic and can cross membranes into the host cell
- NO is also reactive nitrogen species that reacts with superoxide to form a highly toxic oxidant and electron transport inhibitor
DMSP = ROS scavenger and readily crosses membranes
- increased production by host in stress
- Differentially produced by hosts in response to osmotic stress; may be additional antioxidant system
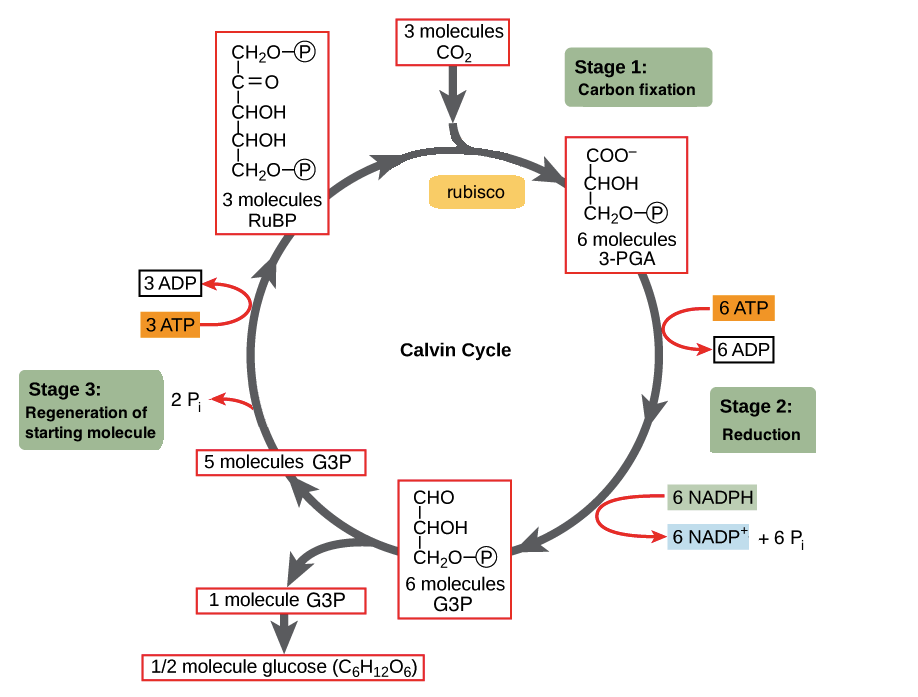
Calvin-Benson Cycle
High demand for DIC from seawater by coral holobiont to support photosynthesis and host calcification.
- DIC in seawater is overwhelmingly bicarbonate (HCO3-) and b/c of its charge cannot cross membranes
- adequate supply of DIC to algal symbionts = critical for productivity and redox state as carbon fixation is ultimate result of electron transport
- b/c of many layers of membranes and high density of symbionts in coral tissues, carbon-concentrating mechanisms are required by both partners
High temps inhibit the Calvin-Benson cycle, limiting carbon fixation as an E sink (not demonstrated in Symbiodiniaceae specifically yet). Photorespiration = fixation of O2 by rubisco, enhanced by high O2:CO2 concentrations. This is not metabolically useful, it is primarily used as an E sink but may consume some abundant O2 and therefore limit ROS production.
Calvin benson cycle:
Symbiosome pathways:

sym
Algal mitochondria result in steady state of ROS production, particularly as H2O2.
Thermal Response of the Coral Host
Chapter 8.3 in the book above.
ROS and reactive nitrogen species are important signaling molecules. Host reaction based on elevated antioxidant activity was seen several days prior to the symbiont’s antioxidant reaction.
- coenzyme Q pool measurement (look into this later..?)
- heat shock response can occur in host without photoinhibition or bleaching
- primary source of ROS in host is the mitochondrian b/c of its role in central metabolism, calcium storage, and apoptosis
Aiptasia host mitochondria undergo morphological changes after thermal shock, followed by mitochondrial permeabilization and degradation, host cell apoptosis, symbiont cell degradation.
Reduced mRNA abundane of cytochrome c and ATP synthase - both central to ETS.
Mitochondria also store calcium, a secondary messenger regulating cell function and sensing cell stress (See Signal-Transduction notes).
- Calcium pool greatly increases, resulting in mitochondrial permeabilization and release of calcium into the cell which triggers the caspase-mediated apoptosis.
- ER is primary store of Ca2+
Calumenin (ER protein) that binds calcium is upregulated in symbiosis, higher temperatures. Particularly in gastrodermal layer
- Calcium-binding proteins may be generally involved in the heat stress response. Seen in A. microphtalma with concurrent higher expression of antioxidant, heat shock, and Ca2+-binding proteins in symbiont population.
Bleaching resistance therefore can depend on the host’s ability to maintain calcium homeostasis.
ER and mitochondria communicate during cellular response. One of ER’s main functions is protein-folding: which is temperature-sensitive and requires the activity of molecular chaperones known as heat shock proteins. Mis-folded proteins trigger the unfolded protein response (UPR): decreased overall protein synthesis, increased protein chaperone and degradation activity, and if stress persists then cell death.
Protein degradation and re-folding in the ER performed by protein disulfide isomerases.
Caspase activity is induced by NO and involved in pro-apoptotic pathways. Apoptosis (programmed cell death) follows a caspase-mediated signaling cascade. Cnidarian apoptosis pathways are highly conserved and more related to vertebrates than other invertebrates.
B-cell lymphoma 2 (Bcl-2): anti-apoptotic protein that mediates mitochondrial membrane permeability; and indirect antioxidant fxn
Mechanisms of symbiont loss
Exocytosis of whole symbiont cells: algal cells are expelled from the host cells in the gastrovascular cavity and out of the mouth of the animal.
Symbiophagy: host autophagic pathways will degrade algal cell within the host cell.
See below figure for the remaining mechanisms.
Symbiont expulsion occurs normally on daily basis as a way to control the symbiont population in a lot of Cnidarians.

fig