3 La especie en paleontología
3.1 El concepto de especie
El concepto de especie es fundamental en biología y existen varios enfoques para definir lo que constituye una especie.
Concepto de Especie Biológica (según Dobzhansky y Mayr): Según este enfoque, una especie es una serie de poblaciones que pueden cruzarse de manera real o potencial para producir descendientes viables y están reproductivamente aisladas de otras poblaciones en condiciones naturales. Este concepto se basa en la capacidad de las poblaciones para intercambiar genes a través de la reproducción sexual y, por lo tanto, se aplica principalmente a organismos que se reproducen sexualmente.
El concepto de especie biológica implica la existencia de barreras reproductivas que pueden ser de naturaleza geográfica o biológica, lo que resulta en la separación de linajes evolutivos. Esta perspectiva considera a las especies como unidades genéticas y evolutivas independientes, libres de seguir su propio curso en respuesta a procesos genéticos y cambios ambientales. Sin embargo, este concepto tiene limitaciones:
Inaplicabilidad en el Registro Fósil: No se puede aplicar el concepto de especie biológica a organismos extintos, ya que no es posible observar directamente su capacidad de reproducción.
Limitaciones en Organismos Asexuales: Este enfoque es limitado en organismos que se reproducen asexualmente, ya que no involucran la reproducción sexual ni las barreras reproductivas.
Hibridación: El concepto de especie biológica no aborda adecuadamente casos de hibridación, donde se producen descendientes fértiles a través del cruce de poblaciones que, en teoría, deberían ser reproductivamente aisladas. Esto es común en plantas vasculares y, en algunos casos, en animales.
Concepto de Especie Ecológica: Según este enfoque, una especie se define como un linaje (o conjunto de linajes cercanamente relacionados) que ocupa una zona adaptativa mínimamente diferente en su distribución en comparación con otras especies. Además, se desarrolla independientemente de todos los linajes establecidos fuera de su área biogeográfica de distribución. Este concepto incorpora nociones de nicho ecológico y exclusión competitiva para explicar las diferencias entre especies basadas en su explotación de recursos ecológicos y adaptación a ambientes específicos.
Concepto de Especie Evolutiva: Este concepto se basa en linajes únicos de poblaciones ancestrales y descendientes que evolucionan independientemente de otros grupos similares. A diferencia del concepto biológico, se centra en el aislamiento genético actual en lugar del potencial. Además, considera que la evolución cladogenética puede ser reticulada, lo que significa que poblaciones previamente separadas pueden reunirse nuevamente, dando lugar a especies híbridas.
Concepto de Especie Filogenética: Según este enfoque, una especie es el grupo monofilético más pequeño con una ascendencia común. Esto se basa en la identificación de un carácter derivado o apomórfico compartido por todos los organismos del grupo. Este enfoque tiene en cuenta similitudes genéticas y moleculares para discernir especies.
Concepto de Especie Morfológica: En este concepto, las especies se definen por diferencias anatómicas medibles. Es especialmente útil en paleontología, donde no siempre se pueden evaluar características de reproducción o genéticas. Sin embargo, este enfoque puede ser limitado, ya que no siempre refleja las diferencias en comportamiento o ecología.
Estos enfoques ofrecen diferentes perspectivas sobre cómo definir y reconocer las especies. En algunos casos, pueden aplicarse a ciertos grupos de organismos y en otros no. La diversidad de conceptos de especies refleja la complejidad de la vida y la necesidad de abordar esta complejidad desde múltiples ángulos para comprender mejor la biodiversidad y la evolución.
Las especies suelen presentar una variabilidad interna, lo que se conoce como politipismo. Este término se utiliza en zoología y botánica para describir grupos taxonómicos que muestran más de un subgrupo en el nivel taxonómico más bajo. En otras palabras, se refiere a la existencia de múltiples formas dentro de una especie, que pueden incluir subespecies y variedades. Dentro de una especie, las subespecies se caracterizan por mantener una uniformidad de tipo, es decir, comparten características distintivas en su apariencia o genética. Además, estas subespecies a menudo ocupan áreas geográficas extensas que son ambientalmente uniformes. Por otro lado, algunas variaciones dentro de una especie pueden estar relacionadas con gradientes ambientales. En lugar de subgrupos claramente definidos, la variación clinal implica que las características de una población cambian gradualmente a medida que te alejas de una ubicación geográfica o entorno específico. Esto refleja la adaptación de la especie a diferentes condiciones ambientales a lo largo de su rango. El politipismo y la variación clinal son fenómenos comunes en la naturaleza y subrayan la adaptabilidad de las especies a una variedad de entornos y condiciones. Estos conceptos son esenciales para comprender la diversidad y la evolución en la biología.
Lectura recomendada
Aldhebiani, A. Y. (2018). Species concept and speciation. Saudi journal of biological sciences, 25(3), 437-440.
De Queiroz, K. (2007). Species concepts and species delimitation. Systematic biology, 56(6), 879-886.
3.2 Especies y especiación
Los mecanismos de aislamiento reproductivo son fundamentales en la especiación, el proceso mediante el cual una población de organismos se divide en dos o más poblaciones reproductivamente aisladas que evolucionan en direcciones diferentes. Estos mecanismos evolutivos evitan que los individuos de diferentes especies se crucen y mezclen sus genes, lo que es esencial para mantener la integridad genética de las especies.
Aislamiento precigótico: Estos mecanismos actúan antes de la fertilización y previenen la formación de híbridos. Son especialmente importantes porque evitan que los recursos y la energía se desperdicien en la producción de descendencia que podría ser inviable o estéril. Los mecanismos de aislamiento precigótico incluyen:
Aislamiento temporal o del hábitat: Este mecanismo opera cuando las poblaciones ocupan diferentes hábitats, lo que impide que los individuos de diferentes especies se encuentren. También puede ser el resultado de barreras físicas, como ríos o montañas. Incluso diferencias en el tiempo de madurez sexual o floración pueden evitar el apareamiento.
Aislamiento conductual: Las especies animales a menudo tienen rituales de apareamiento específicos que son incompatibles entre especies. Estos rituales incluyen comportamientos específicos, señales visuales o auditivas que solo son efectivos dentro de la misma especie.
Aislamiento mecánico: La estructura de los órganos reproductores de las especies puede ser incompatible. Por ejemplo, los genitales de algunos insectos actúan como “cerraduras” y “llaves”, y solo permiten el apareamiento con individuos de la misma especie que tengan estructuras complementarias.
Aislamiento gamético: Incluso si los individuos de diferentes especies se encuentran y copulan, en algunos casos, los óvulos y espermatozoides no pueden fusionarse debido a incompatibilidades gaméticas.
Aislamiento postcigótico: Estos mecanismos operan después de la fertilización y evitan el éxito del cruce entre poblaciones. Pueden resultar en la muerte de los híbridos o en su esterilidad. Algunos de estos mecanismos incluyen:
Mortalidad de cigotos y no viabilidad de híbridos: En algunos cruces entre especies, el cigoto resultante no se desarrolla adecuadamente o es inviable, lo que lleva a la muerte del híbrido. Esto puede deberse a diferencias genéticas importantes entre las especies parentales.
Esterilidad híbrida: En otros casos, los híbridos pueden ser viables pero estériles. Esto ocurre cuando la interacción de los genes de las dos especies resulta en una descendencia que no puede reproducirse. Un ejemplo bien conocido de esto es la mula, un híbrido entre un caballo y una burra.
Estos mecanismos de aislamiento reproductivo son fundamentales en prevenir la hibridación entre especies y mantener la integridad de las poblaciones y la diversidad biológica. Son un ejemplo de cómo la selección natural opera para mantener la separación entre las especies y promover su evolución independiente. Los mecanismos específicos pueden variar entre grupos de organismos, pero todos cumplen la función de garantizar que las poblaciones se mantengan genéticamente distintas.
Los tipos principales de especiación según el origen geográfico de las barreras reproductivas son (Fig. 3.1):
Especiación Alopátrica: La especiación alopátrica se produce cuando las poblaciones se aíslan geográficamente debido a barreras físicas como montañas, ríos u océanos. Este aislamiento impide la reproducción entre las poblaciones, lo que lleva a la acumulación de diferencias genéticas y, finalmente, a la formación de nuevas especies. Un ejemplo clásico es el de los pinzones de Galápagos, que han desarrollado picos adaptados a diferentes fuentes de alimento en islas separadas por el océano.
Especiación Peripátrica: En la especiación peripátrica, una pequeña población se separa de la población principal y queda aislada en un área geográfica limitada. La selección natural y la deriva genética afectan a esta pequeña población de manera diferente, lo que puede resultar en la especiación. Este tipo de especiación se observa a menudo en islas o hábitats aislados donde los grupos pequeños desarrollan características únicas.
Especiación Parapátrica: La especiación parapátrica ocurre cuando las poblaciones tienen rangos geográficos que se superponen parcialmente, pero aún presentan barreras reproductivas debido a diferencias ambientales. Esto provoca que diferentes presiones selectivas actúen en diferentes áreas. Un ejemplo podría ser el de plantas que crecen en suelos contaminados, como el bisonte (Bouteloua dactyloides), que puede tolerar metales pesados, diferenciándose así de otras especies de gramíneas en áreas no contaminadas.
Especiación Simpátrica: La especiación simpátrica ocurre sin una barrera geográfica evidente. En este caso, las poblaciones comparten el mismo espacio, pero desarrollan barreras reproductivas debido a preferencias de hábitat o recursos. Un ejemplo es el mosca de la manzana (Rhagoletis pomonella), que ha comenzado a especializarse en poner huevos en manzanas en lugar de su planta original, el espino, lo que podría llevar a la formación de nuevas especies. Es un concepto controvertido.
Especiación Artificial: La especiación artificial se refiere a la creación de nuevas especies por intervención humana, a menudo a través de experimentos de laboratorio, como en el caso de estudios con drosófilas. Estos experimentos pueden inducir características genéticas específicas que resultan en nuevas especies.
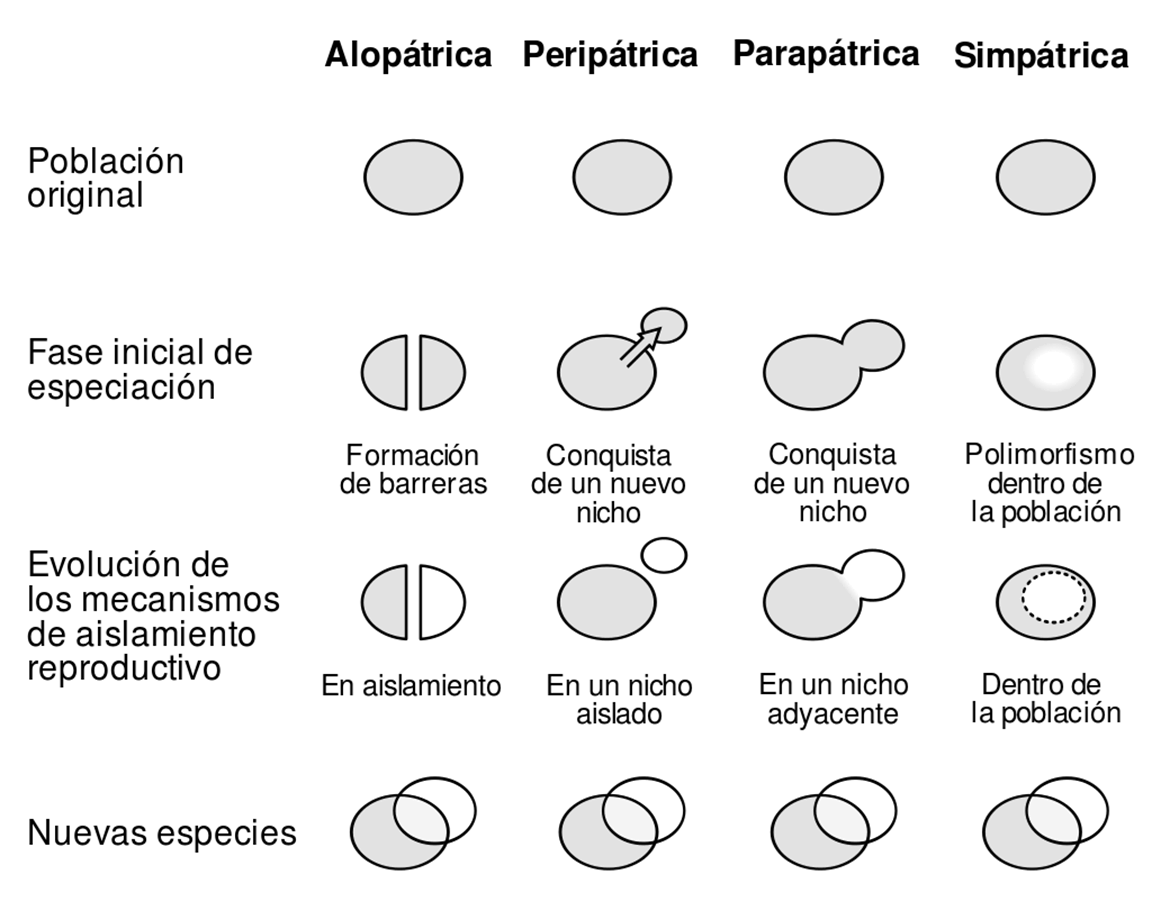
Figura 3.1: Tipos principales de especiación según el origen geográfico de las barreras reproductivas. Imagen disponible bajo licencia Creative Commons CC0.
3.3 Reconocimiento de (morfo)especies en el registro fósil
El reconocimiento de especies en el registro fósil es esencial en paleontología y se basa principalmente en criterios morfológicos. Aunque en la práctica, los zoólogos y botánicos a menudo utilizan evidencia genética para apoyar sus clasificaciones, en muchos fósiles, esta evidencia genética está ausente. Por lo tanto, la taxonomía en paleontología se basa en características morfológicas observables, lo cual puede ser desafiante debido a la naturaleza fragmentaria y a menudo parcial de los fósiles.
Sin embargo, a pesar de estas limitaciones, la discriminación de especies en el registro fósil es posible dentro de un margen de confianza. Esto se debe a la alta correlación que existe entre las características morfológicas de los organismos y otros aspectos esenciales de su biología. De hecho una gran cantidad de trabajos muestra una buena correlación entre la distancia morfológica y genética entre pares de especies, tanto en animales como plantas (Fig. 3.2).
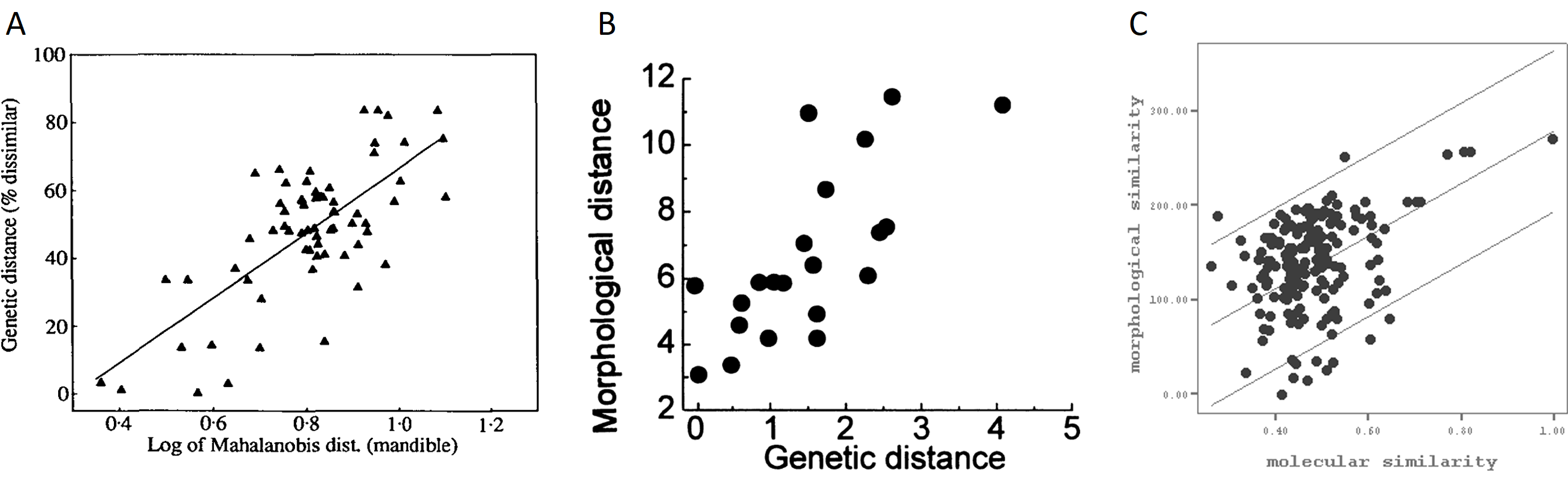
Figura 3.2: Ejemplos de correlación entre distancia morfológica y genética en (A) vertebrados, (B) invertebrados y (C) plantas. Imágenes tomadas de Festing & Roderick (1989), Jackson & Cheetham (1994) y Babić et al. (2012), respectivamente.
En cualquier caso, es obvio que existen ciertas limitaciones en la discriminación de especies en el registro fósil a través de caracteres morfológicos que los paleontólogos deben tener presente:
Caracteres Morfológicos Perdidos: Con el paso del tiempo, es posible que ciertos caracteres morfológicos se pierdan o no se conserven adecuadamente en los fósiles. Esto puede deberse a la degradación de los tejidos blandos o a la erosión de partes del esqueleto. La ausencia de características cruciales puede hacer que la identificación de especies sea más desafiante.
Deformaciones Tafonómicas Sobre la Morfología del Fósil: Durante el proceso de fosilización los fósiles pueden sufrir deformaciones o distorsiones. Esto puede hacer que las características morfológicas de un fósil se vean alteradas, lo que dificulta su correcta identificación y clasificación.
Dimorfismo Sexual: En algunas especies, existe dimorfismo sexual, lo que significa que los individuos de diferentes sexos presentan características morfológicas distintas. En el registro fósil, este fenómeno podría llevar a la identificación errónea de dos especies distintas o a la falta de reconocimiento de una especie dimórfica.
Especies Crípticas: Algunas especies presentan una gran similitud morfológica, lo que dificulta su distinción a través de caracteres visibles. Estas especies crípticas son difíciles de diferenciar solo por sus características morfológicas, lo que puede llevar a una subestimación de la diversidad real en el registro fósil.
Especies Polimórficas: Las especies polimórficas exhiben una variabilidad morfológica entre sus individuos. Esto puede deberse a factores genéticos o ambientales. La variabilidad en la morfología de una especie polimórfica puede llevar a la identificación errónea de diferentes fósiles como especies separadas.
Presencia de Cronoespecies: Las cronoespecies son especies que existieron durante un período de tiempo prolongado y experimentaron cambios morfológicos significativos a lo largo de su historia. Esto puede llevar a la confusión, ya que los fósiles de diferentes etapas de una cronoespecie podrían parecer especies diferentes debido a sus diferencias morfológicas, a pesar de ser parte de la misma especie en evolución.
3.4 (Morfo)especies en el registro fósil y tiempo geológico
3.4.1 Linajes evolutivos
La evolución es un proceso continuo en el que las poblaciones experimentan cambios a lo largo del tiempo debido a factores como la selección natural, la deriva genética y otros mecanismos. En algunos casos, estos cambios son graduales y no generan barreras reproductivas significativas. Sin embargo, en otros, las transformaciones evolutivas pueden ser tan marcadas que impidan o dificulten la hibridación efectiva entre poblaciones de diferentes períodos geológicos.
La presencia de diferencias morfológicas o de registros fósiles notables entre poblaciones en intervalos de tiempo distintos puede indicar que se están tratando con linajes evolutivos separados. Estos linajes pueden clasificarse como especies o subespecies, dependiendo de la magnitud de las diferencias y de la existencia de barreras reproductivas potenciales.
Este enfoque, que considera los linajes en lugar de ceñirse a una única especie, es particularmente relevante al trabajar con el registro fósil, donde la información genética suele ser escasa. Al abordar la evolución en términos de linajes en lugar de limitarse a las especies, los paleontólogos pueden reflejar de manera más precisa la diversidad y la dinámica evolutiva a lo largo del tiempo geológico.
Normalmente, las especies se pueden reconocer durante largos intervalos de tiempo, mostrando variaciones dentro de un rango morfológico estrecho. Sin embargo, surge la pregunta: ¿podrían hibridar la población inicial y la final? Probablemente no, lo que sugiere que no deberíamos considerar estos conjuntos temporales como una única especie. En cambio, es más apropiado hablar de linajes (Fig. 3.3).
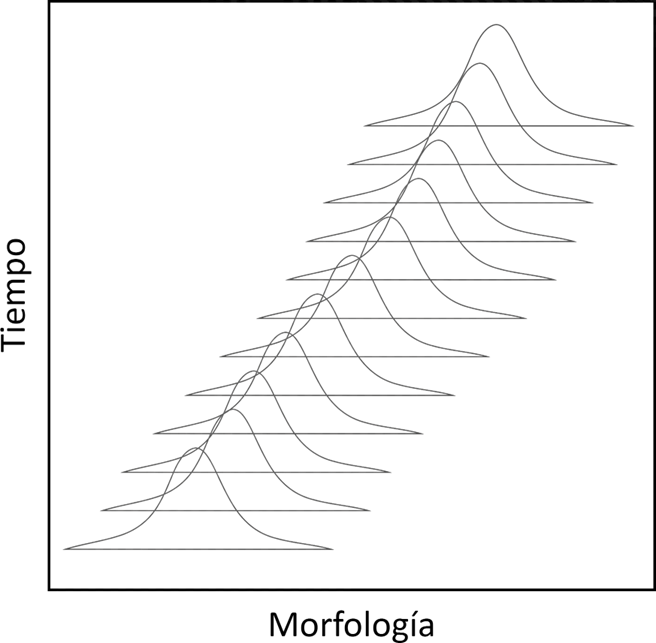
Figura 3.3: Ejemplo gráfico de cómo se documenta el cambio morfológico continuo en una misma población a lo largo del tiempo en el registro fósil.
3.4.2 Plasticidad fenotípica y registro fósil
Es importante destacar que la plasticidad fenotípica, es decir, la capacidad de un organismo para mostrar diferentes rasgos en respuesta a cambios en su entorno, puede llevar a interpretaciones erróneas de cambios morfológicos evolutivos en especies extintas. Esto es particularmente relevante cuando se observa variabilidad coetánea en el registro fósil, lo que significa que diferentes individuos de una misma especie extinta pueden mostrar variaciones morfológicas en un período de tiempo específico.
La variabilidad coetánea puede deberse a la plasticidad fenotípica de una especie, lo que significa que los individuos pueden haber desarrollado diferentes características morfológicas en respuesta a condiciones ambientales cambiantes, pero no necesariamente representa un cambio evolutivo. En otras palabras, los cambios observados en la morfología de los fósiles pueden deberse a una respuesta al su entorno en lugar de cambios genéticos a lo largo de generaciones sucesivas (Fig. 3.4).
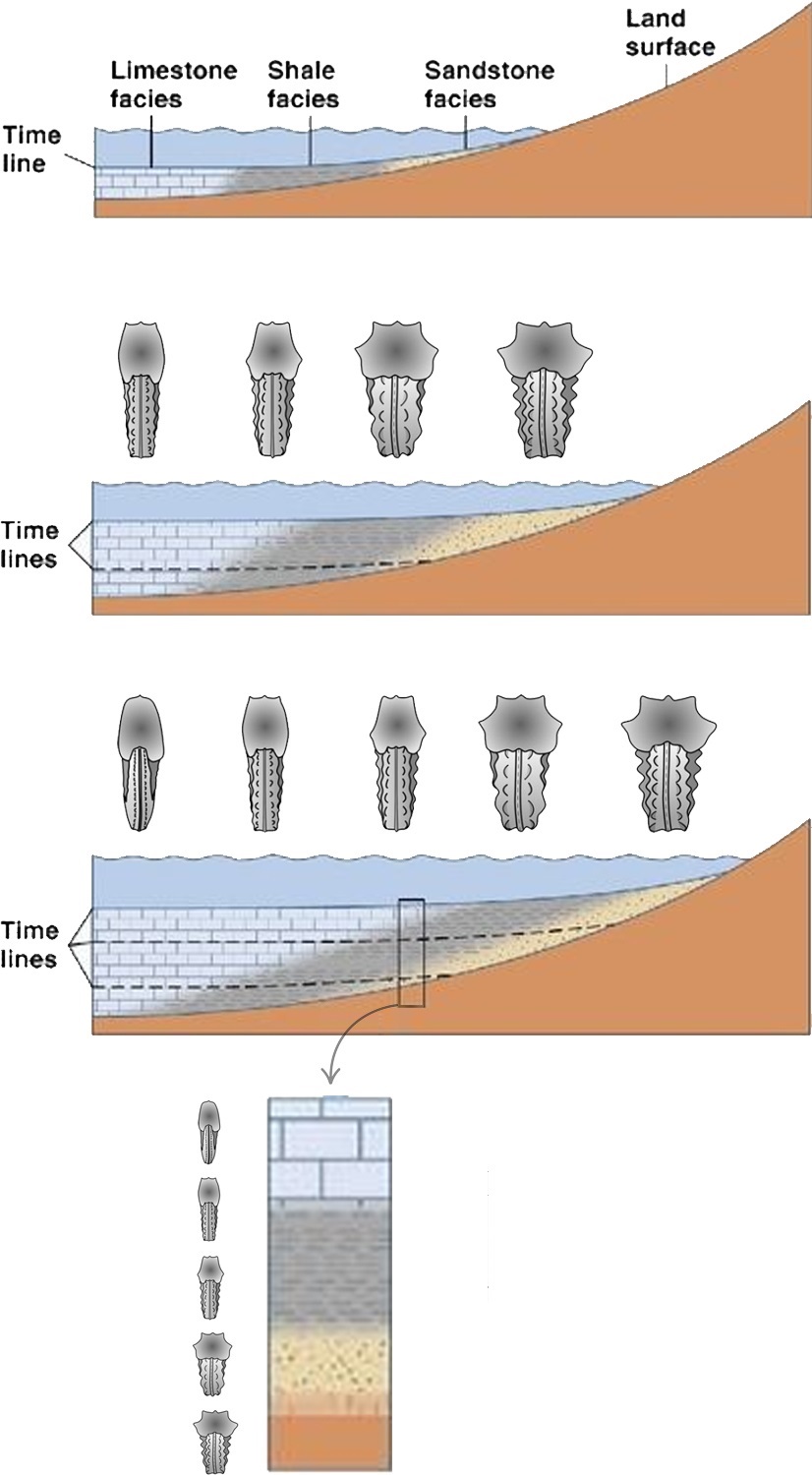
Figura 3.4: Ilustración de un corte transversal de una costa que muestra un proceso de transgresión marina en dirección descendente. En este entorno, las aguas están habitadas por un molusco hipotético que presenta plasticidad fenotípica en su exoesqueleto, variando su morfología en función de la proximidad a la costa. El diagrama inferior representa una columna estratigráfica, en la cual se observa un cambio morfológico a lo largo del tiempo, atribuido exclusivamente a la plasticidad fenotípica de la especie. Imagen modificada de © Geology Desk y Wilmsen & Mosavinia (2011).
Una manera de descartar cambios evolutivos y atribuir la variabilidad morfológica a la plasticidad fenotípica es buscar correlaciones entre los diferentes morfotipos y las facies o los tipos de entorno en los que se encuentran los fósiles. Esta correlacion puede buscarse entre niveles sincronicos o a lo alrgo de series temporales (Fig. 3.5). Si se encuentra dicha correlación entre las características morfológicas y los entornos específicos, esto puede sugerir que la variabilidad morfológica es el resultado de plasticidad fenotípica. Sin embargo, no siempre es posible establecer esta correlación, y en tales casos, la distinción entre plasticidad fenotípica y cambio evolutivo puede ser más desafiante. Por lo tanto, los paleontólogos deben utilizar múltiples líneas de evidencia y ser conscientes de la plasticidad fenotípica al interpretar cambios morfológicos en especies extintas.
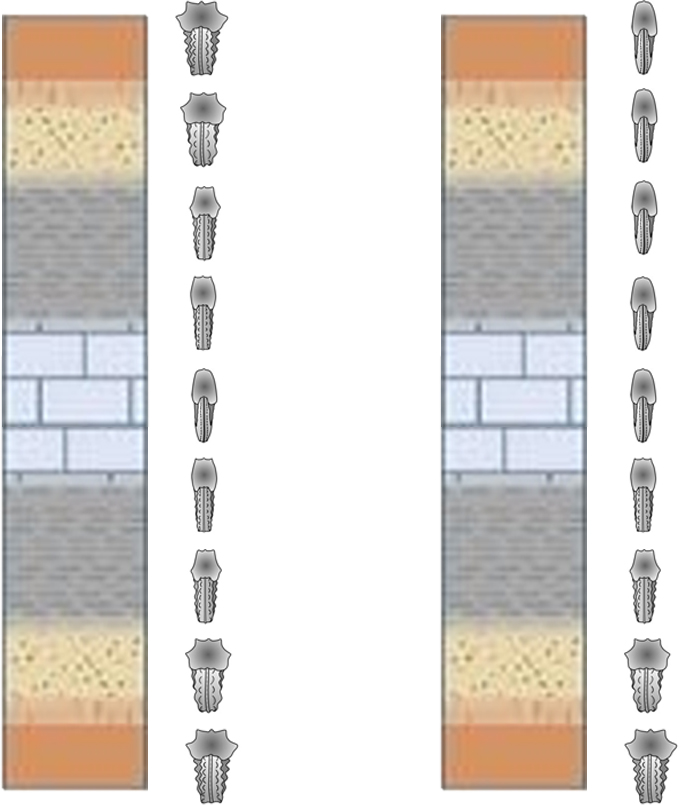
Figura 3.5: Ilustración de dos columnas estratigráficas donde, la de la izquierda muestra una clara correspondencia entre facies y morfotipos, lo que sugiere que los cambios morfológicos en este caso pueden atribuirse a la plasticidad fenotípica. En contraste, la columna de la derecha no refleja tal relación, indicando cambio evolutivo. Imagen modificada de © Geology Desk y Wilmsen & Mosavinia (2011).
Lectura recomendada
Lister, A. M. (2021). Phenotypic plasticity in the fossil record. In Phenotypic Plasticity & Evolution (pp. 267-297). CRC Press.